Bacteriile sunt cel mai vechi grup de organisme existente în prezent pe Pământ. Primele bacterii au apărut probabil cu mai bine de 3,5 miliarde de ani în urmă și timp de aproape un miliard de ani au fost singurele creaturi vii de pe planeta noastră. Întrucât aceștia au fost primii reprezentanți ai naturii vii, corpul lor avea o structură primitivă.
De-a lungul timpului, structura lor a devenit mai complexă, dar până în prezent bacteriile sunt considerate cele mai primitive organisme unicelulare. Este interesant că unele bacterii păstrează încă trăsăturile primitive ale strămoșilor lor antici. Acest lucru se observă la bacteriile care trăiesc în izvoarele fierbinți cu sulf și nămolul anoxic de la fundul rezervoarelor.
Majoritatea bacteriilor sunt incolore. Doar câteva sunt violete sau verzi. Dar coloniile multor bacterii au o culoare strălucitoare, care este cauzată de eliberarea unei substanțe colorate în mediu sau de pigmentarea celulelor.
Descoperitorul lumii bacteriilor a fost Antony Leeuwenhoek, un naturalist olandez al secolului al XVII-lea, care a creat pentru prima dată un microscop de mărire perfect care mărește obiectele de 160-270 de ori.
Bacteriile sunt clasificate ca procariote și sunt clasificate într-un regn separat - Bacteriile.
Forma corpului
Bacteriile sunt organisme numeroase și diverse. Ele variază ca formă.
| Numele bacteriei | Forma bacteriilor | Imaginea bacteriilor |
| Cocci | În formă de minge | |
| Bacil |  | În formă de tijă |
| Vibrio | În formă de virgulă | |
| Spirillum |  | Spirală |
| Streptococi |  | Lanț de coci |
| Stafilococ |  | Ciorchini de coci |
| Diplococul | Două bacterii rotunde închise într-o capsulă mucoasă |
Metode de transport
Printre bacterii există forme mobile și imobile. Motile se deplasează datorită contracțiilor în formă de undă sau cu ajutorul flagelilor (fire elicoidale răsucite), care constau dintr-o proteină specială numită flagelină. Pot exista unul sau mai mulți flageli. La unele bacterii sunt situate la un capăt al celulei, în altele - la două sau pe toată suprafața.

Dar mișcarea este, de asemenea, inerentă multor alte bacterii care nu au flageli. Astfel, bacteriile acoperite la exterior cu mucus sunt capabile să alunece.
Unele bacterii acvatice și din sol lipsite de flageli au vacuole gazoase în citoplasmă. Într-o celulă pot exista 40-60 de vacuole. Fiecare dintre ele este umplut cu gaz (probabil azot). Prin reglarea cantității de gaz din vacuole, bacteriile acvatice se pot scufunda în coloana de apă sau se pot ridica la suprafața acesteia, iar bacteriile din sol se pot deplasa în capilarele solului.
Habitat
Datorită simplității lor de organizare și lipsei de pretenții, bacteriile sunt răspândite în natură. Bacteriile se găsesc peste tot: într-o picătură chiar și din cea mai pură apă de izvor, în boabele de sol, în aer, pe stânci, în zăpada polară, nisipurile deșertului, pe fundul oceanului, în uleiul extras din adâncimi mari și chiar în apa din izvoarele termale cu o temperatura de aproximativ 80ºC. Ei trăiesc pe plante, fructe, diverse animale și la oameni în intestine, cavitatea bucală, membre și pe suprafața corpului.
Bacteriile sunt cele mai mici și mai numeroase creaturi vii. Datorită dimensiunilor lor mici, pătrund cu ușurință în orice fisuri, crăpături sau pori. Foarte rezistent și adaptat la diverse condiții de viață. Tolerează uscarea, frigul extrem și încălzirea până la 90 ° C fără a-și pierde viabilitatea.
Practic nu există niciun loc pe Pământ unde bacteriile să nu se găsească, dar în cantități variate. Condițiile de viață ale bacteriilor sunt variate. Unele dintre ele necesită oxigen atmosferic, altele nu au nevoie de el și sunt capabile să trăiască într-un mediu fără oxigen.
În aer: bacteriile se ridică în atmosfera superioară până la 30 km. și altele.
Există mai ales multe dintre ele în sol. 1 g de sol poate conține sute de milioane de bacterii.
În apă: în straturile de suprafață ale apei din rezervoare deschise. Bacteriile acvatice benefice mineralizează reziduurile organice.
În organismele vii: bacteriile patogene pătrund în organism din mediul extern, dar numai în condiții favorabile provoacă boli. Simbioticele trăiesc în organele digestive, ajutând la descompunerea și absorbția alimentelor și la sintetizarea vitaminelor.
Structura externă
Celula bacteriană este acoperită cu o înveliș special dens - un perete celular, care îndeplinește funcții de protecție și de susținere și, de asemenea, conferă bacteriei o formă permanentă, caracteristică. Peretele celular al unei bacterii seamănă cu peretele unei celule vegetale. Este permeabil: prin ea, nutrienții trec liber în celulă, iar produsele metabolice ies în mediu. Adesea, bacteriile produc un strat protector suplimentar de mucus deasupra peretelui celular - o capsulă. Grosimea capsulei poate fi de multe ori mai mare decât diametrul celulei în sine, dar poate fi și foarte mică. Capsula nu este o parte esențială a celulei; se formează în funcție de condițiile în care se află bacteriile. Protejează bacteriile de uscare.
Pe suprafața unor bacterii există flageli lungi (unul, doi sau mai mulți) sau vilozități scurte și subțiri. Lungimea flagelului poate fi de multe ori mai mare decât dimensiunea corpului bacteriei. Bacteriile se mișcă cu ajutorul flagelilor și vilozităților.

Structura interna
În interiorul celulei bacteriene există citoplasmă densă, imobilă. Are o structură stratificată, nu există vacuole, prin urmare diverse proteine (enzime) și nutrienți de rezervă sunt localizate în substanța citoplasmei în sine. Celulele bacteriene nu au nucleu. O substanță care transportă informații ereditare este concentrată în partea centrală a celulei lor. Bacterii, - acid nucleic - ADN. Dar această substanță nu este formată într-un nucleu.
Organizarea internă a unei celule bacteriene este complexă și are propriile sale caracteristici specifice. Citoplasma este separată de peretele celular prin membrana citoplasmatică. În citoplasmă există o substanță principală, sau matrice, ribozomi și un număr mic de structuri membranare care îndeplinesc o varietate de funcții (analogi ai mitocondriilor, reticulului endoplasmatic, aparatul Golgi). Citoplasma celulelor bacteriene conține adesea granule de diferite forme și dimensiuni. Granulele pot fi compuse din compuși care servesc ca sursă de energie și carbon. Picături de grăsime se găsesc și în celula bacteriană.
În partea centrală a celulei este localizată substanța nucleară - ADN, care nu este delimitat de citoplasmă de o membrană. Acesta este un analog al nucleului - un nucleoid. Nucleoidul nu are o membrană, un nucleol sau un set de cromozomi.
Metode de alimentație
Bacteriile au diferite metode de hrănire. Printre aceștia se numără autotrofe și heterotrofe. Autotrofele sunt organisme care sunt capabile să producă în mod independent substanțe organice pentru nutriția lor.
Plantele au nevoie de azot, dar nu pot absorbi singure azotul din aer. Unele bacterii combină moleculele de azot din aer cu alte molecule, rezultând substanțe care sunt disponibile plantelor.
Aceste bacterii se instalează în celulele rădăcinilor tinere, ceea ce duce la formarea unor îngroșări pe rădăcini, numite noduli. Astfel de noduli se formează pe rădăcinile plantelor din familia leguminoaselor și ale altor plante.

Rădăcinile furnizează carbohidrați bacteriilor, iar bacteriile rădăcinilor furnizează substanțe care conțin azot care pot fi absorbite de plantă. Coabitarea lor este reciproc avantajoasă.
Rădăcinile plantelor secretă o mulțime de substanțe organice (zaharuri, aminoacizi și altele) cu care bacteriile se hrănesc. Prin urmare, în special multe bacterii se instalează în stratul de sol din jurul rădăcinilor. Aceste bacterii transformă resturile de plante moarte în substanțe disponibile pentru plante. Acest strat de sol se numește rizosferă.
Există mai multe ipoteze despre pătrunderea bacteriilor nodulare în țesutul radicular:
- prin deteriorarea țesutului epidermic și a cortexului;
- prin firele de păr din rădăcină;
- numai prin membrana celulară tânără;
- datorită bacteriilor însoțitoare care produc enzime pectinolitice;
- datorită stimulării sintezei acidului B-indoleacetic din triptofan, prezent întotdeauna în secrețiile rădăcinilor plantelor.
Procesul de introducere a bacteriilor nodulare în țesutul radicular constă în două faze:
- infecția firelor de păr din rădăcină;
- procesul de formare a nodulilor.
În cele mai multe cazuri, celula invadatoare se înmulțește activ, formează așa-numitele fire de infecție și, sub forma unor astfel de fire, se deplasează în țesutul plantei. Bacteriile nodulare care ies din firul de infecție continuă să se înmulțească în țesutul gazdă.

Celulele vegetale pline cu celule care se înmulțesc rapid de bacterii nodulare încep să se dividă rapid. Conectarea unui nodul tânăr cu rădăcina unei plante leguminoase se realizează datorită fasciculelor vascular-fibroase. În timpul perioadei de funcționare, nodulii sunt de obicei denși. În momentul în care apare activitatea optimă, nodulii capătă o culoare roz (mulțumită pigmentului de leghemoglobină). Numai acele bacterii care conțin leghemoglobină sunt capabile să fixeze azotul.
Bacteriile nodulare creează zeci și sute de kilograme de îngrășământ cu azot pe hectar de sol.
Metabolism
Bacteriile diferă unele de altele prin metabolismul lor. În unele apare cu participarea oxigenului, în altele - fără el.
Majoritatea bacteriilor se hrănesc cu substanțe organice gata preparate. Doar câteva dintre ele (albastru-verde sau cianobacteriile) sunt capabile să creeze substanțe organice din cele anorganice. Ele au jucat un rol important în acumularea de oxigen în atmosfera Pământului.
Bacteriile absorb substanțele din exterior, își rup moleculele în bucăți, își adună învelișul din aceste părți și își reumple conținutul (așa cresc) și aruncă moleculele inutile. Învelișul și membrana bacteriei îi permit să absoarbă numai substanțele necesare.

Dacă învelișul și membrana unei bacterii ar fi complet impermeabile, nicio substanță nu ar intra în celulă. Dacă ar fi permeabile la toate substanțele, conținutul celulei s-ar amesteca cu mediul - soluția în care trăiește bacteria. Pentru a supraviețui, bacteriile au nevoie de o înveliș care să permită trecerea substanțelor necesare, dar nu a substanțelor inutile.
Bacteria absoarbe nutrienții aflați în apropierea ei. Ce se întâmplă mai departe? Dacă se poate mișca independent (prin mișcarea unui flagel sau împingând mucusul înapoi), atunci se mișcă până când găsește substanțele necesare.
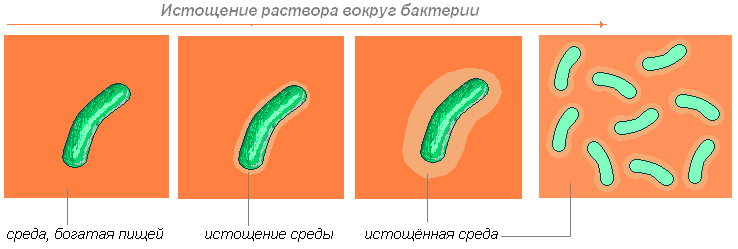
Dacă nu se poate mișca, atunci așteaptă până când difuzia (capacitatea moleculelor unei substanțe de a pătrunde în desișul de molecule ale unei alte substanțe) aduce moleculele necesare.
Bacteriile, împreună cu alte grupuri de microorganisme, efectuează o muncă chimică enormă. Prin transformarea diferiților compuși, ei primesc energia și nutrienții necesari vieții lor. Procesele metabolice, metodele de obținere a energiei și nevoia de materiale pentru construirea substanțelor corpului lor sunt diverse în bacterii.
Alte bacterii își satisfac toate nevoile de carbon necesare sintezei substanțelor organice în organism în detrimentul compușilor anorganici. Se numesc autotrofi. Bacteriile autotrofe sunt capabile să sintetizeze substanțe organice din cele anorganice. Printre acestea se numără:
Chemosinteza
Utilizarea energiei radiante este cea mai importantă, dar nu singura modalitate de a crea materie organică din dioxid de carbon și apă. Sunt cunoscute bacterii care nu folosesc lumina solară ca sursă de energie pentru o astfel de sinteză, ci energia legăturilor chimice care apar în celulele organismelor în timpul oxidării anumitor compuși anorganici - hidrogen sulfurat, sulf, amoniac, hidrogen, acid azotic, compuși feroși de fier și mangan. Ei folosesc materia organică formată folosind această energie chimică pentru a construi celulele corpului lor. Prin urmare, acest proces se numește chimiosinteză.
Cel mai important grup de microorganisme chemosintetice sunt bacteriile nitrificatoare. Aceste bacterii trăiesc în sol și oxidează amoniacul format în timpul descompunerii reziduurilor organice în acid azotic. Acesta din urmă reacționează cu compușii minerali ai solului, transformându-se în săruri ale acidului azotic. Acest proces are loc în două etape.
Bacteriile de fier transformă fierul feros în fier oxid. Hidroxidul de fier rezultat se depune și formează așa-numitul minereu de fier din mlaștină.
Unele microorganisme există datorită oxidării hidrogenului molecular, oferind astfel o metodă autotrofă de nutriție.
O trăsătură caracteristică a bacteriilor cu hidrogen este capacitatea de a trece la un stil de viață heterotrof atunci când sunt furnizate cu compuși organici și absența hidrogenului.
Astfel, chimioautotrofele sunt autotrofe tipice, deoarece sintetizează în mod independent compușii organici necesari din substanțe anorganice și nu îi iau gata preparati din alte organisme, cum ar fi heterotrofele. Bacteriile chimioautotrofe diferă de plantele fototrofe prin independența lor completă față de lumină ca sursă de energie.
Fotosinteza bacteriană
Unele bacterii cu sulf care conțin pigment (violet, verde), care conțin pigmenți specifici - bacterioclorofilele, sunt capabile să absoarbă energia solară, cu ajutorul căreia hidrogenul sulfurat din corpurile lor este descompus și eliberează atomi de hidrogen pentru a reface compușii corespunzători. Acest proces are multe în comun cu fotosinteza și diferă doar prin aceea că, la bacteriile violet și verzi, donatorul de hidrogen este hidrogen sulfurat (ocazional acizi carboxilici), iar la plantele verzi este apa. În ambele, separarea și transferul hidrogenului se realizează datorită energiei razelor solare absorbite.
Această fotosinteză bacteriană, care are loc fără eliberarea de oxigen, se numește fotoreducere. Fotoreducerea dioxidului de carbon este asociată cu transferul de hidrogen nu din apă, ci din hidrogen sulfurat:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Semnificația biologică a chimiosintezei și fotosintezei bacteriene la scară planetară este relativ mică. Doar bacteriile chemosintetice joacă un rol semnificativ în procesul de ciclizare a sulfului în natură. Absorbit de plantele verzi sub formă de săruri de acid sulfuric, sulful este redus și devine parte a moleculelor proteice. În plus, atunci când rămășițele de plante și animale moarte sunt distruse de bacteriile putrefactive, sulful este eliberat sub formă de hidrogen sulfurat, care este oxidat de bacteriile sulfuroase la sulf liber (sau acid sulfuric), formând în sol sulfiți accesibili plantelor. Bacteriile chimio- și fotoautotrofe sunt esențiale în ciclul azotului și sulfului.
Sporularea
Sporii se formează în interiorul celulei bacteriene. În timpul procesului de sporulare, celula bacteriană suferă o serie de procese biochimice. Cantitatea de apă liberă din el scade și activitatea enzimatică scade. Acest lucru asigură rezistența sporilor la condiții de mediu nefavorabile (temperatură ridicată, concentrație mare de sare, uscare etc.). Sporularea este caracteristică doar unui grup mic de bacterii.

Sporii sunt o etapă opțională în ciclul de viață al bacteriilor. Sporularea începe doar cu lipsa nutrienților sau acumularea de produse metabolice. Bacteriile sub formă de spori pot rămâne latente mult timp. Sporii bacterieni pot rezista la fierbere prelungită și la înghețare foarte lungă. Când apar condiții favorabile, sporul germinează și devine viabil. Sporii bacterieni sunt o adaptare pentru a supraviețui în condiții nefavorabile.
Reproducere
Bacteriile se reproduc prin împărțirea unei celule în două. După ce a atins o anumită dimensiune, bacteria se împarte în două bacterii identice. Apoi fiecare dintre ei începe să se hrănească, crește, se împarte și așa mai departe.

După alungirea celulei, se formează treptat un sept transversal, apoi celulele fiice se separă; În multe bacterii, în anumite condiții, după divizare, celulele rămân conectate în grupuri caracteristice. În acest caz, în funcție de direcția planului de diviziune și de numărul de diviziuni, apar diferite forme. Reproducerea prin înmugurire are loc ca o excepție la bacterii.
În condiții favorabile, diviziunea celulară în multe bacterii are loc la fiecare 20-30 de minute. Cu o reproducere atât de rapidă, descendenții unei bacterii în 5 zile sunt capabili să formeze o masă care poate umple toate mările și oceanele. Un calcul simplu arată că se pot forma 72 de generații (720.000.000.000.000.000.000 de celule) pe zi. Dacă este convertit în greutate - 4720 de tone. Cu toate acestea, acest lucru nu se întâmplă în natură, deoarece majoritatea bacteriilor mor rapid sub influența razelor solare, uscării, lipsei hranei, încălzirii la 65-100 ° C, ca urmare a luptei dintre specii etc.
Bacteria (1), după ce a absorbit suficientă hrană, crește în dimensiune (2) și începe să se pregătească pentru reproducere (diviziunea celulară). ADN-ul său (într-o bacterie molecula de ADN este închisă într-un inel) se dublează (bacteria produce o copie a acestei molecule). Ambele molecule de ADN (3,4) se găsesc atașate de peretele bacteriei și, pe măsură ce bacteria se alungește, se depărtează (5,6). Mai întâi nucleotida se divide, apoi citoplasma.

După divergența a două molecule de ADN, apare o constricție pe bacterie, care împarte treptat corpul bacteriei în două părți, fiecare conținând o moleculă de ADN (7).
Se întâmplă (în Bacillus subtilis) să se lipească două bacterii și să se formeze o punte între ele (1,2).

Jumperul transportă ADN-ul de la o bacterie la alta (3). Odată ajunse într-o singură bacterie, moleculele de ADN se împletesc, se lipesc împreună în unele locuri (4) și apoi schimbă secțiuni (5).
Rolul bacteriilor în natură
Gyre
Bacteriile sunt cea mai importantă verigă în ciclul general al substanțelor din natură. Plantele creează substanțe organice complexe din dioxid de carbon, apă și săruri minerale din sol. Aceste substanțe revin în sol cu ciuperci moarte, plante și cadavre de animale. Bacteriile descompun substanțele complexe în unele simple, care sunt apoi folosite de plante.
Bacteriile distrug substanțele organice complexe ale plantelor moarte și cadavrele animalelor, excrețiile organismelor vii și diverse deșeuri. Hranindu-se cu aceste substante organice, bacteriile saprofite de degradare le transforma in humus. Acestea sunt un fel de ordonanți ai planetei noastre. Astfel, bacteriile participă activ la ciclul substanțelor din natură.
Formarea solului
Deoarece bacteriile sunt distribuite aproape peste tot și apar în număr mare, ele determină în mare măsură diferite procese care au loc în natură. Toamna, frunzele copacilor și arbuștilor cad, mor lăstarii supraterane de ierburi, cad ramuri bătrâne și din când în când trunchiurile copacilor bătrâni cad. Toate acestea se transformă treptat în humus. În 1 cm3. Stratul de suprafață al solului forestier conține sute de milioane de bacterii saprofite ale solului din mai multe specii. Aceste bacterii transformă humusul în diferite minerale care pot fi absorbite din sol de rădăcinile plantelor.
Unele bacterii din sol sunt capabile să absoarbă azotul din aer, folosindu-l în procesele vitale. Aceste bacterii fixatoare de azot trăiesc independent sau se stabilesc în rădăcinile plantelor de leguminoase. După ce au pătruns în rădăcinile leguminoaselor, aceste bacterii provoacă creșterea celulelor radiculare și formarea de noduli pe ele.
Aceste bacterii produc compuși de azot pe care îi folosesc plantele. Bacteriile obțin carbohidrați și săruri minerale din plante. Astfel, există o relație strânsă între planta de leguminoase și bacteriile nodulare, care este benefică atât pentru unul cât și pentru celălalt organism. Acest fenomen se numește simbioză.
Datorită simbiozei cu bacteriile nodulare, plantele leguminoase îmbogățesc solul cu azot, ajutând la creșterea randamentului.
Distribuția în natură
Microorganismele sunt omniprezente. Singurele excepții sunt craterele vulcanilor activi și zonele mici de la epicentrele bombelor atomice explodate. Nici temperaturile scăzute din Antarctica, nici fluxurile de fierbere ale gheizerelor, nici soluțiile saturate de sare din bazinele de sare, nici insolația puternică a vârfurilor muntilor, nici iradierea dură a reactoarelor nucleare nu interferează cu existența și dezvoltarea microflorei. Toate ființele vii interacționează constant cu microorganismele, fiind adesea nu doar depozitele lor, ci și distribuitorii lor. Microorganismele sunt native ale planetei noastre, explorând activ cele mai incredibile substraturi naturale.
Microflora solului
Numărul de bacterii din sol este extrem de mare - sute de milioane și miliarde de indivizi pe gram. Sunt mult mai mulți în sol decât în apă și aer. Numărul total de bacterii din sol se modifică. Numărul de bacterii depinde de tipul de sol, de starea acestora și de adâncimea straturilor.
Pe suprafața particulelor de sol, microorganismele sunt localizate în microcolonii mici (20-100 de celule fiecare). Ele se dezvoltă adesea în grosimea cheagurilor de materie organică, pe rădăcinile plantelor vii și pe moarte, în capilare subțiri și în interiorul bulgări.
Microflora solului este foarte diversă. Aici există diferite grupe fiziologice de bacterii: bacterii putrefactoare, bacterii nitrificante, bacterii fixatoare de azot, bacterii sulfuroase etc. printre ele se numără aerobe și anaerobe, forme spori și non-spori. Microflora este unul dintre factorii de formare a solului.
Zona de dezvoltare a microorganismelor în sol este zona adiacentă rădăcinilor plantelor vii. Se numește rizosferă, iar totalitatea microorganismelor conținute în ea se numește microfloră rizosferă.
Microflora rezervoarelor
Apa este un mediu natural în care microorganismele se dezvoltă în număr mare. Cea mai mare parte a acestora intră în apa din sol. Un factor care determină numărul de bacterii din apă și prezența nutrienților în aceasta. Cele mai curate ape sunt din fântâni și izvoare arteziene. Rezervoarele deschise și râurile sunt foarte bogate în bacterii. Cel mai mare număr de bacterii se găsește în straturile de suprafață ale apei, mai aproape de țărm. Pe măsură ce vă îndepărtați de țărm și creșteți în adâncime, numărul bacteriilor scade.
Apa curată conține 100-200 de bacterii pe ml, iar apa poluată conține 100-300 de mii sau mai mult. Există multe bacterii în nămolul de jos, în special în stratul de suprafață, unde bacteriile formează o peliculă. Acest film conține o mulțime de bacterii cu sulf și fier, care oxidează hidrogenul sulfurat în acid sulfuric și, prin urmare, împiedică moartea peștilor. Există mai multe forme purtătoare de spori în nămol, în timp ce formele care nu poartă spori predomină în apă.
În ceea ce privește compoziția speciilor, microflora apei este similară cu microflora solului, dar există și forme specifice. Prin distrugerea diferitelor deșeuri care intră în apă, microorganismele realizează treptat așa-numita purificare biologică a apei.
Microflora aerului
Microflora aerului este mai puțin numeroasă decât microflora solului și a apei. Bacteriile se ridică în aer cu praf, pot rămâne acolo o perioadă de timp, apoi se așează pe suprafața pământului și mor din cauza lipsei de nutriție sau sub influența razelor ultraviolete. Numărul de microorganisme din aer depinde de zona geografică, teren, perioada anului, poluarea cu praf etc. fiecare fir de praf este un purtător de microorganisme. Majoritatea bacteriilor sunt în aer deasupra întreprinderilor industriale. Aerul din mediul rural este mai curat. Cel mai curat aer este deasupra pădurilor, munților și zonelor înzăpezite. Straturile superioare de aer conțin mai puțini microbi. Microflora aerului conține multe bacterii pigmentate și purtătoare de spori, care sunt mai rezistente decât altele la razele ultraviolete.
Microflora corpului uman
Corpul uman, chiar și unul complet sănătos, este întotdeauna un purtător de microfloră. Când corpul uman intră în contact cu aerul și solul, diverse microorganisme, inclusiv cele patogene (bacili tetanici, gangrena gazoasă etc.), se instalează pe îmbrăcăminte și pe piele. Cele mai frecvent expuse părți ale corpului uman sunt contaminate. E. coli și stafilococii se găsesc pe mâini. Există peste 100 de tipuri de microbi în cavitatea bucală. Gura, cu temperatura, umiditatea și reziduurile sale nutritive, este un mediu excelent pentru dezvoltarea microorganismelor.
Stomacul are o reacție acidă, astfel încât majoritatea microorganismelor din el mor. Pornind de la intestinul subțire, reacția devine alcalină, adică. favorabil microbilor. Microflora din intestinul gros este foarte diversă. Fiecare adult excretă zilnic aproximativ 18 miliarde de bacterii în excremente, adică. mai mulți indivizi decât oameni de pe glob.
Organele interne care nu sunt conectate la mediul extern (creier, inimă, ficat, vezică urinară etc.) sunt de obicei lipsite de microbi. Microbii intră în aceste organe numai în timpul bolii.
Bacteriile din ciclul substanțelor
Microorganismele în general și bacteriile în special joacă un rol important în ciclurile importante din punct de vedere biologic ale substanțelor de pe Pământ, efectuând transformări chimice care sunt complet inaccesibile fie plantelor, fie animalelor. Diferite etape ale ciclului elementelor sunt efectuate de organisme de diferite tipuri. Existența fiecărui grup individual de organisme depinde de transformarea chimică a elementelor efectuată de alte grupuri.
Ciclul azotului
Transformarea ciclică a compușilor azotați joacă un rol primordial în furnizarea formelor necesare de azot organismelor biosferei cu nevoi nutriționale diferite. Peste 90% din fixarea totală a azotului se datorează activității metabolice a anumitor bacterii.
Ciclul carbonului
Transformarea biologică a carbonului organic în dioxid de carbon, însoțită de reducerea oxigenului molecular, necesită activitatea metabolică comună a diferitelor microorganisme. Multe bacterii aerobe efectuează oxidarea completă a substanțelor organice. În condiții aerobe, compușii organici sunt descompuși inițial prin fermentație, iar produșii organici finali ai fermentației sunt oxidați în continuare prin respirație anaerobă dacă sunt prezenți acceptori anorganici de hidrogen (nitrat, sulfat sau CO2).
Ciclul sulfului
Sulful este disponibil organismelor vii în principal sub formă de sulfați solubili sau compuși organici redusi de sulf.
Ciclul fierului
Unele corpuri de apă dulce conțin concentrații mari de săruri reduse de fier. În astfel de locuri, se dezvoltă o microfloră bacteriană specifică - bacterii de fier, care oxidează fierul redus. Ei participă la formarea minereurilor de fier din mlaștină și a surselor de apă bogate în săruri de fier.
Bacteriile sunt cele mai vechi organisme, apărând în urmă cu aproximativ 3,5 miliarde de ani în Arhean. Timp de aproximativ 2,5 miliarde de ani au dominat Pământul, formând biosfera și au participat la formarea atmosferei de oxigen.
Bacteriile sunt una dintre cele mai simplu organisme vii structurate (cu excepția virusurilor). Se crede că acestea sunt primele organisme care au apărut pe Pământ.
Bacteriile sunt microorganisme procariote, al căror material genetic este reprezentat în principal de un singur ADN circular dublu catenar, numit cromozom de geneticieni. În cazuri relativ rare, un cromozom este reprezentat de o moleculă de ADN liniară.
Dimensiunea acestui ADN este mult mai mare decât dimensiunea celulei bacteriene în sine. Deci, de exemplu, la E coli lungimea ADN-ului cromozomial este de 1300 µm (1,3 mm - 4,6 x 10 6 bp), iar dimensiunea celulei este de 1,1-1,5 x 2,0-6,0 µm. Mai mult decât atât, ADN-ul nu umple întreaga celulă, ci este conținut doar într-o zonă limitată, constituind, foarte aproximativ, o treime din volumul celulei.
   |
Fig.1. Genomul bacterian și diagrama nivelurilor sale de compactare.
Rezultă că ADN-ul există într-o celulă într-o stare foarte ordonată (condensată) sub forma unei structuri compacte. Această structură, care amintește vag de nucleele eucariote, se numește nucleoidși este vizibilă la microscop numai după colorații specifice ADN-ului (Fig. 1). Într-un microscop electronic, apare ca o formațiune constând din numeroase bucle care se extind dintr-o regiune centrală densă. Formarea unui număr mare (până la 140 pe genom) de bucle numite domenii, este unul dintre nivelurile de compactare a ADN-ului. Fiecare domeniu este ancorat la bază de o moleculă de ARN și constă din aproximativ 40 kb. Buclele de ADN nu sunt sub forma unui duplex alungit liber, ci au un al doilea nivel de compactare datorită răsucirii în formațiuni superhelical prin conexiunea cu proteinele HLP.
Aceste proteine sunt de dimensiuni mici, foarte bazice și se leagă puternic de ADN. În compoziția de aminoacizi, ele seamănă cu histonele eucariote.
Nucleoidul nu este separat de citoplasmă prin membrana nucleară și este atașat de mezosomi– invaginări specifice ale membranei citoplasmatice în celulă. Legarea ADN-ului de o regiune specifică a membranei este necesară pentru funcționarea genomului.
Molecula circulară de ADN a bacteriilor (cromozomul) este o moleculă genetică auto-replicabilă - replicon. Replicarea începe cu puncte de inițiere a replicării (ori– original ) , localizat, de regulă, în locul atașării ADN-ului la membrană. Din punctul de inițiere, replicarea are loc secvenţial, bidirecţional, folosind un mecanism semi-conservator. Replicarea se termină la regiune de terminare a replicarii (ter), situat pe o secțiune de ADN circular opus originii replicării (Fig. 2).
Orez. 3. Distribuția copiilor fiice ale ADN-ului și diviziunea celulară bacteriană.
Numărul de cromozomi dintr-o celulă bacteriană depinde de stadiul de dezvoltare și de condițiile fiziologice de creștere a culturii. În stadiul de creștere logaritmică E coli per 1 nucleoid există 2,8 echivalenți ADN ai unui genom, datorită segregării lente a doi cromozomi fiice sau reinițialării unor noi cicluri de replicare a ADN-ului chiar înainte de diviziunea celulară (Fig. 4).
 |
Fig.4. Numărul de cromozomi dintr-o celulă în stadiile staționare (A) și logaritmice (B) ale creșterii culturii.
În unele bacterii, celulele conțin în mod normal nu unul, ci mulți cromozomi. Pot forma unul sau mai mulți nucleoizi. Există, de asemenea, o dependență a conținutului de ADN dintr-o celulă de dimensiunea acesteia, deși aceasta nu înseamnă o modificare corespunzătoare a volumului de informații genetice.
ADN-ul bacterian este caracterizat printr-o densitate mare a genelor (1 genă la 1 kb). ADN-ul care codifică proteine reprezintă aproximativ 85-90% din tot ADN-ul. Dimensiunea medie a secvențelor de ADN dintre gene este de numai 110-125 bp. ADN-ul bacterian necodant reprezintă mai puțin de 1% și este de obicei prezent sub formă de transpozoni. Deci, în ADN-ul tulpinii Escherichia coli Linia K12 MG 1655 a găsit 41 de copii ale diverșilor transpozoni (IS), care sunt implicați în procesele de inserție și excludere a plasmidelor. Mulți fagi, deși nu sunt complet excluși din genomul bacterian, lasă o parte din genele lor acolo ca o urmă. Aceste rămășițe, incapabile de mișcare și dezvoltare independentă, sunt numite fagi „criptici”.
Intronii sunt extrem de rari în genomul bacterian. Există cazuri de suprapunere a genelor, în care o genă este situată în interiorul alteia pe aceeași catenă de ADN. Genomul bacterian este caracterizat prin operoni: E coli 27% dintre unitățile de transcripție prezise sunt operoni.
O celulă bacteriană poate conține alți repliconi care pot exista separat de cromozomul bacterian. Ei sunt numiti, cunoscuti plasmide. Plasmidele sunt molecule circulare (la unele specii liniare) de ADN dublu catenar de diferite dimensiuni de la 1000 bp. până la aproape o treime din dimensiunea cromozomului bacterian însuși. Numărul și gama de plasmide din celulele bacteriene pot varia. Diferențele în spectrul plasmidelor sunt adesea observate chiar și între celulele diferitelor tulpini ale aceleiași specii bacteriene. Unele plasmide pot fi inserate în cromozomul bacterian, făcând parte din repliconul bacterian și pot fi excluse din acesta din nou, restabilind forma unui replicon autonom. Astfel de plasmide se numesc epizomi.
Profagele pot fi incluse și în materialul genetic al bacteriilor.
Virușii care conțin ADN fie au propriile enzime de replicare (în capsidă), fie genomul lor codifică informații despre sinteza enzimelor virale care asigură replicarea acidului nucleic viral. Cantitatea acestor enzime variază atunci când este aplicată la diferiți viruși. De exemplu, genomul virusului bacterian T4 codifică informații despre sinteza a aproximativ 30 de enzime virale. În plus, genomul virusurilor mari codifică nucleaze care distrug ADN-ul celulei gazdă, precum și proteine, al căror efect asupra ARN polimerazei celulare este însoțit de faptul că „ARN polimeraza tratată în acest mod transcrie diferite gene virale. în diferite stadii de infecție virală. În schimb, virusurile ADN mici sunt mai dependente de enzimele celulei gazdă. De exemplu, sinteza ADN-ului adenovirusurilor este asigurată de enzimele celulare.[...]
ADN-ul bacterian este un compus cu înaltă polimeri format dintr-un număr mare de nucleotide - polinucleotide cu o greutate moleculară de aproximativ 4 milioane.O moleculă de ADN este un lanț de nucleotide, în care aranjamentul lor are o anumită secvență. Secvența bazelor azotate codifică informația genetică a fiecărei specii. Încălcarea acestei secvențe este posibilă din cauza mutațiilor naturale sau sub influența factorilor mutageni. În acest caz, microorganismul câștigă sau pierde unele proprietăți. Caracteristicile lui se schimbă ereditar, adică apare o nouă formă a microorganismului. În toate microorganismele - procariote și eucariote - purtătorii de informații genetice sunt acizii nucleici - ADN și ARN. Doar unii viruși sunt o excepție: nu au ADN, iar informațiile ereditare sunt înregistrate sau reflectate doar în ARN.[...]
În celulele bacteriene, numărul total de baze ADN conține 32-65 mol.% guanină și citozină.[...]
Nucleul unei celule bacteriene. Aproximativ 1-2% din masa uscată a microorganismelor este ADN, care conține informațiile genetice ale organismului. Majoritatea microorganismelor au o zonă (sau mai multe zone) în care se concentrează cea mai mare parte a ADN-ului, având o structură specifică (sau organel) și numită nucleu. Nucleul (sau substanța nucleară) este asociat cu o membrană citoplasmatică, indiferent dacă este înconjurat de membrane elementare (ca la amibe) sau fără ele (ca la bacterii și alge albastru-verzi). Substanța nucleară este activată în timpul perioadei de reproducere și odată cu debutul modificărilor legate de vârstă asociate cu îmbătrânirea celulară.[...]
Segmentul de ADN (gena) care este destinat clonării moleculare trebuie să aibă capacitatea de a se replica atunci când este transferat într-o celulă bacteriană, adică să fie un replicon. Cu toate acestea, el nu are o astfel de abilitate. Prin urmare, pentru a asigura transferul și detectarea genelor clonate în celule, acestea sunt combinate cu așa-numiții vectori genetici. Acesta din urmă trebuie să aibă cel puțin două proprietăți. În primul rând, vectorii trebuie să fie capabili de a se replica în celule și în mai multe copii. În al doilea rând, ele trebuie să ofere capacitatea de a selecta celulele care conțin vectorul, adică să aibă un marker care poate fi utilizat pentru a contra-selecta celulele care conțin vectorul împreună cu gena donată (molecule de ADN recombinant). Plasmidele și fagii îndeplinesc aceste cerințe. Plasmidele sunt vectori buni deoarece sunt repliconi și pot conține gene de rezistență la orice antibiotic, ceea ce permite selectarea bacteriilor pentru rezistența la acest antibiotic și, prin urmare, detectarea ușoară a moleculelor de ADN recombinant. [...]
În bacterii, ADN-ul este împachetat mai puțin strâns, spre deosebire de nucleele adevărate; Un nucleoid nu are o membrană, un nucleol sau un set de cromozomi. ADN-ul bacterian nu este asociat cu principalele proteine - histonele - și este localizat în nucleoid sub forma unui mănunchi de fibrile.[...]
Utilizarea tehnicilor ADN recombinant pentru a produce agenți biologici pentru controlul poluării este în stadii incipiente, dar o tehnică care se poate dovedi utilă în viitorul apropiat este sondarea genetică. Selecția organismelor capabile să transforme un nou compus se bazează adesea pe capacitatea de a utiliza substanța ca substrat de creștere. Dacă creșterea este slabă sau substratul este doar metabolizat, atunci metodele de selecție vor fi nepotrivite pentru identificarea capacității de degradare. Prin urmare, ar fi util să se dezvolte sondaje genetice pentru a identifica secvențe specifice în plasmide și cromozomi, acest lucru fiind necesar pentru a determina potențialul catabolic, chiar dacă acest potențial nu este exprimat. Astfel de sonde sunt proiectate pentru plasmidele TOL. Metoda poate identifica o colonie bacteriană care conține o plasmidă TOL printre 106 colonii de Escherichia coli. Un astfel de instrument puternic va fi de mare valoare în izolarea funcțiilor catabolice ascunse.[...]
Dezvoltarea unei tehnici elegante pentru „clonarea” ADN-ului pentru a produce un număr mare de copii exacte ale fragmentelor specifice de ADN (Fig. 13.4) a deschis recent noi orizonturi în studiul structurii, organizării și funcției genomului. Dacă scindați DIC dublu catenar cu una dintre enzimele de „restricție” (una dintre nucleaze), care recunosc și scindează în mod specific secvențe scurte de nucleotide (4-6 perechi), atunci apar fragmente de ADN foarte reproductibile. Capetele a două catene de ADN sunt de obicei deplasate unul față de celălalt datorită specificității locurilor de tăiere ale unei molecule dublu catenare, ale cărei catene sunt complementare în compoziția de bază. ADN-ul este de obicei inserat într-o genă plasmidă importantă pentru reproducere, cum ar fi o genă de rezistență la antibiotice, care permite bacteriilor care conțin plasmida să crească în prezența antibioticului.[...]
În bacterii, replicarea produce multe copii ale plasmidelor și, în acest fel, cantități mari de fragmente de ADN încorporate pot fi „crescute” și apoi pur și simplu izolate din nou prin digestie cu aceeași enzimă de restricție și separarea produselor rezultate prin electroforeză pe gel. Utilizarea acestei metode de recombinare a ADN-ului a revoluționat studiul genelor.[...]
S-a descoperit recent că razele de intensitate scăzută cu o lungime de undă de 320-400 nm (o regiune apropiată de zona luminii vizibile) au un efect mutagen asupra virusurilor ADN bacteriene. Posibilul efect al radiațiilor în acest interval de lungimi de undă asupra virusurilor plantelor nu a fost încă descoperit.[...]
Curbele dependenței de reasociere față de COT obținute pentru ADN-ul bacterian sunt lipsite de îndoituri, iar DIC ale eucariotelor se reasociază după un tip diferit (Fig. 13.2). La concentrații scăzute de ADN și un timp scurt de incubare, o proporție semnificativă de ADN monocatenar este renaturalizată, iar odată cu creșterea COT, se formează o cantitate suplimentară de molecule dublu catenare, astfel încât se obține o curbă în două faze. . Reaturarea rapidă la valori scăzute ale COT arată că unele secvențe la eucariote se repetă de multe ori, adică de până la 10.000 de ori sau mai mult. [...]
Absența CXC poate fi imitată și în cazurile în care ADN-ul fagilor de testare nu conține situsuri recunoscute de enzima de restricție existentă în tulpina studiată. Acest fenomen reprezintă una dintre variantele modificărilor adaptative evolutive ale virusurilor bacteriene menite să îi ajute să depășească bariera CXC. Efectul presiunii de selecție în acest caz particular este exprimat într-o scădere semnificativă statistic a numărului sau chiar eliminarea completă în ADN-ul fag a secvențelor de nucleotide care sunt substratul enzimelor de restricție caracteristice celulelor gazdă ale virusului bacterian.[.. .]
Lindegren a descris etapele posibile ale formării unui bacteriofag din ADN-ul profag, sugerând că profagul ia naștere ca un fragment de ADN bacterian străin care a intrat accidental în celulă, care în stadiile incipiente se împarte sincron cu ADN-ul bacterian. Următoarea etapă importantă în dezvoltarea virusului ar fi o astfel de schimbare a profagului, în urma căreia reproducerea sa, independentă de ADN-ul celulei gazdă, a devenit posibilă; ca urmare, profagul va fi folosit. toate nucleotidele disponibile, perturbând astfel creșterea celulei gazdă. În cele din urmă, într-o etapă ulterioară, s-ar putea forma o înveliș proteic de protecție și ar apărea alte proteine, care ar asigura supraviețuirea ADN-ului în afara corpului gazdă și infecția eficientă a celulelor noi. Fragmentul separat de ADN bacterian a codificat inițial proteine adaptate funcțiilor bacteriene. Modificări foarte semnificative ale ADN-ului sunt necesare pentru a apărea obiecte la fel de complexe și specializate precum, de exemplu, fagul T2 al E. coli, care conțin și baze care sunt absente în DIC bacterian. [...]
Informația genetică a bacteriilor nu se limitează la ADN-ul situat în nucleoidul celulei bacteriene. După cum sa menționat deja în secțiunile anterioare ale cărții, elementele extracromozomiale, numite colectiv plasmide, servesc și ca purtători de proprietăți ereditare. Spre deosebire de echivalentele nucleare ADN, nucleoizii, care sunt organitele unei celule bacteriene, plasmidele sunt elemente genetice independente. Pierderea plasmidelor sau achiziția lor nu afectează biologia celulei (achiziția plasmidelor are un efect pozitiv doar asupra populației în ansamblu, crescând viabilitatea speciei). Plasmidele transmisibile le includ pe cele care inițiază proprietățile donorului în celulele gazdă. În același timp, acestea din urmă primesc o nouă calitate - capacitatea de a se conjuga cu celulele primitoare și de a le oferi plasmidele lor. Celulele primitoare, care dobândesc plasmide în timpul conjugării, se transformă ele însele în donatori.[...]
Absența adsorbției nu epuizează varietatea de opțiuni de interacțiune dintre virusurile bacteriene și celulele microbiene. Ele ilustrează doar o latură a acestui fenomen, și anume manifestarea mecanismelor de protecție celulară care fenotipic (după criteriul lipsei de creștere) imită restricția. Cu toate acestea, există o altă variantă a interacțiunii celulă-bacteriofag care poate imita absența CXC. Exemple de astfel de mecanisme sunt sinteza inhibitorilor și metilazelor codificate de genele fagice care protejează ADN-ul viral de acțiunea enzimelor de restricție de tip II.[...]
Mecanismul efectului dezinfectant al clorului este asociat cu tulburările metabolice ale celulei bacteriene în timpul procesului de dezinfecție a apei. În același timp, a fost evidențiat un efect asupra activității enzimatice a bacteriilor, în special asupra dehidrogenazelor care catalizează reacțiile redox în celula bacteriană. A. M. Skidalskaya (1969) a studiat efectul clorului asupra procesului de decarboxilare a aminoacizilor bacterieni, care are loc în prezența enzimelor decarboxilază strict specifice și, de asemenea, a determinat compoziția nucleotidică a ADN-ului Escherichia coli după finalizarea procesului de dezinfecție la diferite niveluri. cu efect bactericid.[ . ..]
Bacteriofagii din grupul T au forma unor bețișoare care măsoară 100 x 25 nm. Genomul lor este reprezentat de ADN. Sunt fagi virulenți, deoarece după ce infectează celulele bacteriene, acestea din urmă sunt lizate, eliberând un număr mare de particule de fagi nou sintetizate. [...]
Plasmidele bacteriene sunt structuri genetice situate în citoplasmă și reprezentând molecule de ADN cu dimensiuni cuprinse între 2250 și 400.000 de perechi de baze azotate. Ele există separat de cromozomi în cantități de la una la câteva zeci de copii per celulă bacteriană.[...]
Tulpina Pseu.dom.onas vug1 ae ri. pka8eoIco1a are o plasmidă lungă de 150 mii bp, care se poate replica autonom sau poate fi integrată în cromozomul bacterian. Excizia imprecisă ulterioară a produs o familie de plasmide cu lungimea cuprinsă între 35 și 270 kb, dintre care unele conțineau segmente mari de ADN cromozomial.[...]
În timpul evoluției, bacteriile au dezvoltat capacitatea de a sintetiza așa-numitele enzime de restricție (endonucleaze), care au devenit parte a sistemului celular (bacterian) de restricție-modificare. La bacterii, sistemele de restricție-modificare sunt un sistem imunitar intracelular pentru protejarea împotriva ADN-ului străin. Spre deosebire de organismele superioare, în care recunoașterea și distrugerea virusurilor, bacteriilor și a altor agenți patogeni are loc extracelular, în bacterii, protecția împotriva ADN-ului străin (ADN-ul plantelor și animalelor în ale căror corpuri trăiesc) are loc intracelular, adică atunci când ADN-ul străin pătrunde în citoplasmă. de bacterii. Pentru a se proteja, bacteriile au dezvoltat și capacitatea de a-și „eticheta” propriul ADN cu baze de metilare pe anumite secvențe. Din acest motiv, ADN-ul străin, din cauza absenței grupărilor metil pe aceleași secvențe, este topit (tăiat) în fragmente de către diferite enzime de restricție bacteriene, iar apoi degradat de exonucleazele bacteriene în nucleotide. Putem spune că în acest fel bacteriile se protejează de ADN-ul plantelor și animalelor, în corpurile cărora trăiesc temporar (ca agenți patogeni) sau permanent (ca saprofite).[...]
Proprietățile ereditare ale bacteriilor sau caracteristicile individuale sunt codificate în unități de ereditate - gene, localizate liniar în cromozom de-a lungul catenei de ADN. În consecință, o genă este un fragment dintr-o catenă de ADN.Fiecărei caracteristici îi corespunde o anumită genă și, adesea, unei părți și mai mici de ADN - un codon. Cu alte cuvinte, catena de ADN conține informații despre toate proprietățile bacteriilor într-o ordine liniară. Cu toate acestea, bacteriile au încă o caracteristică. Nucleele eucariotelor conțin de obicei mai mulți cromozomi, numărul acestora în nucleu este constant la fiecare specie. Nucleoidul bacterian conține doar un inel al unei catene de ADN, adică un cromozom. Cu toate acestea, suma caracteristicilor ereditare ale unei celule bacteriene nu este epuizată de stocul de informații conținut într-un cromozom sau într-o spirală de ADN dublu catenară închisă în formă de inel. Plasmidele conțin ADN, care transportă și informații genetice transmise de la celula mamă la celula fiică.[...]
Mutațiile sunt modificări ale aparatului genic al unei celule, care sunt însoțite de modificări ale caracteristicilor controlate de aceste gene. Există macro și microdaune la ADN, ceea ce duce la modificări ale proprietăților celulei. Macro-modificări, și anume: pierderea unei secțiuni a ADN-ului (diviziunea), mișcarea unei secțiuni separate (translocarea) sau rotația unei anumite secțiuni a moleculei cu 180° (inversare) sunt observate relativ rar la bacterii.Microleziuni sau mutații punctuale. , adică sunt mult mai tipice pentru ei, modificări calitative ale genelor individuale, de exemplu, înlocuirea unei perechi de baze azotate. Mutațiile pot fi directe și inverse sau inverse. Mutațiile directe sunt mutații în organismele de tip sălbatic, de exemplu, pierderea capacității de a sintetiza în mod independent factorii de creștere, adică o tranziție de la proto- la auxotrofie. Mutațiile din spate reprezintă o întoarcere sau o întoarcere la tipul sălbatic. Capacitatea de a reveni este caracteristică mutațiilor punctuale. Ca urmare a mutațiilor, se modifică caracteristici atât de importante precum capacitatea de a sintetiza independent aminoacizi și vitamine (mutanți auxotrofici) și capacitatea de a forma enzime. Aceste mutații sunt numite biochimice. Mutațiile care duc la modificări ale sensibilității la antibiotice și alte substanțe antimicrobiene sunt de asemenea bine cunoscute. Pe baza originii lor, mutațiile sunt împărțite în spontane și induse. Spontanele apar spontan, fără intervenția umană și sunt aleatorii în natură. Frecvența unor astfel de mutații este foarte scăzută și variază de la 1 X 10"4 la 1 X 10-10. Cele induse apar atunci când microorganismele sunt expuse la factori mutageni fizici sau chimici. Factorii fizici care au un efect mutagen includ radiațiile ultraviolete și ionizante, precum și temperatura. O serie de compuși sunt mutageni chimici, iar dintre aceștia cei mai activi sunt așa-numiții supermutageni. În condiții naturale și în experimente, pot apărea modificări în compoziția populațiilor bacteriene ca urmare a acțiunii a doi factori - mutații și autoselecție, care apare ca urmare a adaptării unor mutanți la condițiile de mediu. Acest proces se observă în mod evident într-un mediu în care sursa alimentară predominantă este o substanță sintetică, de exemplu, un surfactant sau caprolactamă.[...]
O singură celulă de E. coli este înconjurată de o membrană celulară cu trei straturi de aproximativ 40 nm grosime, care este o „pungă” sau „plic” care conține conținut celular sub formă de aproximativ 2 x 10 16 g de proteine, 6 x 10 16 g de ADN și 2 x 10 14 g de ARN (în principal ARN ribozomal). Într-o celulă bacteriană sunt sintetizate aproximativ 2000 de proteine diferite, dintre care majoritatea se găsesc în citoplasmă. Concentrația unor proteine este de 10“® M, în timp ce altele sunt de ordinul a 2 x 10“4 M (de la 10 la 200.000 de molecule per celulă).[...]
În organismele unicelulare, reproducerea sexuală are loc sub mai multe forme. Conjugarea se găsește și la ciliați, în care în timpul acestui proces are loc un transfer de nuclee de la un individ la altul, urmat de divizarea acestuia din urmă.[...]
Bacterii: organisme unicelulare procariote (“prenucleare”). Celulele lor nu au un nucleu separat de citoplasmă. Cu toate acestea, programul genetic, ca și cel al tuturor organismelor vii, este codificat ca o secvență de nucleotide în ADN și poartă informații despre structura proteinelor. Celulele bacteriene nu conțin organele precum cloroplastele (specializate pentru fotosinteză) și mitocondriile (specializate pentru respirația celulară și sinteza ATP). Aceste procese biochimice apar în bacterii din citoplasmă.[...]
Dimensiunile celulelor extrem de mici sunt o caracteristică, dar nu și principala caracteristică a bacteriilor. Toate bacteriile sunt reprezentate de un tip special de celulă care nu are un nucleu adevărat înconjurat de o membrană nucleară. Un analog al nucleului din bacterii este nucleoidul - plasmă care conține ADN, nedelimitat de citoplasmă printr-o membrană. În plus, celulele bacteriene se caracterizează prin absența mitocondriilor, a cloroplastelor, precum și prin structura și compoziția specială a structurilor membranare și a pereților celulari. Organismele ale căror celule nu au un nucleu adevărat sunt numite procariote (prenucleare) sau protocite (adică organisme cu o organizare celulară primitivă).[...]
Celulele de micoplasmă au formă ovală și dimensiunea lor este de aproximativ 0,1-0,25 nm în diametru (Fig. 43). Ele se caracterizează prin prezența unei membrane plasmatice exterioare subțiri (grosime - aproximativ 8 nm), care înconjoară citoplasma care conține o moleculă de ADN suficientă pentru a codifica aproximativ 800 de proteine diferite, ARN de diferite tipuri, ribozomi cu un diametru de aproximativ 20 nm. Citoplasma lor conține diverse incluziuni sub formă de proteine, granule lipidice și alți compuși. Din cauza rigidității celulare insuficiente, membranele de micoplasmă trec prin filtre bacteriene.[...]
S-a stabilit că pe ribozomi are loc legarea aminoacizilor activați și plasarea lor într-un lanț polipeptidic în conformitate cu informațiile genetice primite de la nucleu prin ARN-ul mesager (ARNm), care citește informația corespunzătoare din ADN și o transmite către ribozomii. Pe ribozomi izolați sunt sintetizate un număr de proteine și se remarcă includerea în ei a aminoacizilor marcați. Rolul matricei în sinteza proteinelor este îndeplinit de ARNm, care este atașat de ribozom. La suprafața acestuia din urmă are loc o interacțiune între un complex de aminoacizi, ARN de transfer care poartă următorul aminoacid și secvența de nucleotide a ARN mesager, care funcționează pe ribozom o singură dată și, după sinteza lanțului polipeptidic, se dezintegrează. , iar proteina nou sintetizată se acumulează în ribozomi. Într-o celulă bacteriană, cu o perioadă de regenerare de 90 de minute, rata de turnover a ARNm ajunge la 4-6 secunde.[...]
Citoplasma este o soluție coloidală, a cărei fază dispersată este compuși proteici complexi și substanțe asemănătoare grăsimilor, iar mediul de dispersie este apa. Unele forme de bacterii conțin incluziuni în citoplasmă - picături de grăsime, sulf, glicogen etc. Componentele permanente ale celulelor bacteriene sunt excrescențe speciale ale membranei citoplasmatice - mezosomi, care conțin sisteme redox enzimatice. În aceste formațiuni au loc procese asociate în principal cu respirația bacteriilor. În incluziuni mici - ribozomi care conțin acid ribonucleic, are loc biosinteza proteinelor. Cele mai multe tipuri de bacterii nu au un nucleu separat. Substanța nucleară, reprezentată de ADN, nu este separată de citoplasmă și formează un nucleoid. Transportul substanțelor necesare vieții celulei și îndepărtarea produselor metabolice se realizează prin canale și cavități speciale, separate de citoplasmă printr-o membrană având aceeași structură cu cea citoplasmatică. Această formațiune structurală se numește reticul endoplasmatic (reticul).[...]
O idee despre variabilitatea și ereditatea bacteriilor nu se poate forma fără cunoașterea unor prevederi ale geneticii moleculare a celulei procariote. Procesele de adaptare a culturilor microbiene la condițiile de mediu în schimbare se bazează pe variabilitate și ereditate, care sunt secțiuni ale geneticii bacteriene. La prezentarea citologiei unei celule bacteriene, structura ADN-ului și ARN-ului și rolul lor în viața celulei au fost deja luate în considerare. Structura caracteristică a ADN-ului este păstrată la fiecare specie și este transmisă descendenților din generație în generație, ca și alte caracteristici. ADN-ul bacterian este un helix dublu catenar care se închide într-un inel. Catena inelata a ADN-ului bacterian, situata in nucleoid, nu contine proteine. Acest inel ADN corespunde cromozomului unei celule eucariote. Se știe că cromozomul celulelor eucariote, pe lângă ADN, conține întotdeauna o componentă proteică. Rezultă că conceptul de cromozom la eucariote este oarecum diferit de conceptul de cromozom bacterian. Catena de ADN care alcatuieste cromozomul bacterian, desigur, difera de la specie la specie. Componenta fosfat de zahăr a ADN-ului este aceeași în toate tipurile de bacterii; aranjarea bazelor azotate și combinarea lor, dimpotrivă, diferă între diferitele specii.[...]
Utilizarea crescândă fără discernământ a antibioticelor la animale, care sunt utilizate în doze mici ca promotori de creștere și, de asemenea, ca măsură preventivă împotriva tulburărilor gastrointestinale legate de stres la animalele de fermă, duce la creșterea prevalenței rezistenței la antibiotice a factorului R în populațiile microbiene. , transmis de la o celulă bacteriană la alta în timpul conjugării. Transferul are loc printr-o plasmidă, care este un ADN circular extracromozomial capabil de replicare.[...]
Spre deosebire de fagii virulenți, sunt cunoscuți așa-numiții fagi cu acțiune moderată, sau pur și simplu fagi moderati. Un reprezentant tipic al unor astfel de fagi este fagul X, care a fost și este folosit ca model experimental pentru a clarifica multe probleme ale geneticii moleculare. Fagul X are două proprietăți importante. La fel ca fagii virulenți, poate infecta celulele bacteriene, se poate reproduce vegetativ, producând sute de copii în celule și poate liza celulele eliberând particule de fagi mature. Cu toate acestea, ADN-ul acestui fag poate fi inclus în cromozomul bacterian, transformându-se într-un profag. În acest caz, are loc așa-numita lizogenizare a bacteriilor, iar bacteriile care conțin profagul sunt numite lizogenice. Celulele bacteriene lizogenice pot poseda un profag pentru o perioadă nedeterminată de timp fără a fi lizate. Liza cu eliberarea de noi particule de fagi este observată după expunerea bacteriilor lizogenice la orice factor, de exemplu, radiația UV, care induce dezvoltarea unui profag într-un fag. Studiul bacteriilor lizogenice a făcut posibilă obținerea unui număr de date noi despre rolul diferitelor proteine în acțiunea genelor fagilor.[...]
Genomul de cloroplast al unui număr de plante superioare este format din 120 de gene. Genomul cloroplastului este foarte asemănător cu genomul bacterian atât în ceea ce privește organizarea, cât și funcționalitatea. Genomul mitocondrial uman nu are probabil introni, dar intronii se găsesc în ADN-ul cloroplastelor unor plante superioare, precum și în ADN-ul mitocondriilor ciupercilor. Se crede că genomul de cloroplast al plantelor superioare rămâne neschimbat timp de aproximativ câteva milioane de ani. Este posibil ca o astfel de antichitate să fie și caracteristică genomului mitocondrial al mamiferelor, inclusiv al oamenilor.[...]
Diagramele moderne care ilustrează activitatea genelor sunt construite pe baza unei analize logice a datelor experimentale obținute prin metode biochimice și genetice. Utilizarea unor metode subtile de microscopie electronică permite să vedem literalmente activitatea aparatului ereditar al celulei. Recent, au fost obținute imagini microscopice electronice care arată cum pe matricea ADN bacteriană, în acele zone în care moleculele de ARN polimerază (o enzimă care catalizează transcripția ADN-ului în ARN) sunt atașate de ADN, are loc sinteza moleculelor de ARNm. . Catenele de ARNm, situate perpendicular pe molecula liniară de ADN, se deplasează de-a lungul matricei și cresc în lungime. Pe măsură ce catenele de ARN se prelungesc, ribozomii sunt atașați de ele, care, la rândul lor, deplasându-se de-a lungul catenei de ARN către ADN, duc la sinteza proteinelor.[...]
Transducția este transferul de material genetic de la o bacterie donatoare la o bacterie primitoare folosind un fag. Fenomenul de transducție a fost descoperit pentru prima dată în 1951 de către Lederberg și colegii săi în Salmonella typhimurium. În zilele noastre se face o distincție între transducția nespecifică și cea specifică. Cu transducție nespecifică, fagul poate transfera orice trăsătură de la o bacterie donatoare la o bacterie primitoare. Transferul se realizează numai de către fagi temperați (non-virulenți). Fagii temperați sunt capabili să infecteze bacteriile, dar nu se reproduc în ele și nu provoacă liză, ci sunt incluși în ADN-ul celulei bacteriene și în această stare neinfecțioasă sub forma unui așa-numit profag sunt transmise din celulă. la celulă în timpul reproducerii. Culturile bacteriene care conțin un profag sunt numite lizogenice. În aceste culturi, cu o frecvență scăzută (în una din 102 - 105 celule), se observă reproducerea spontană a fagului și liza celulară are loc cu eliberarea de particule de fagi, detectate cu ajutorul bacteriilor indicator pentru care un astfel de fag este virulent. . [...]
Experimentele au fost efectuate pe o celulă cu trei camere, constând dintr-o cameră centrală de lucru și două camere cu electrozi. 750 mg de vată au fost plasate într-o cameră de lucru de 25 X 7 X 37 mm (lungime X lățime X înălțime), separată de camerele electrozilor prin membrane de celofan. Prin intermediul acesteia, soluția inițială a substanțelor studiate a fost alimentată de jos în sus cu o viteză constantă. Conținutul de compuși în soluțiile inițiale furnizate în camera de lucru (C0) și în soluțiile care părăsesc camera (Ci) a fost monitorizat prin maximele de absorbție a proteinelor și acizilor nucleici în intervalul de număr de undă (35,5-38) X 103 cm- 1 folosind un spectrofotometru Specord UV-VIS UV. Camerele electrozilor au fost umplute cu cărbune activ granular și apă distilată a fost trecută prin ele într-un flux separat.
Principalul secret al vieții organice constă în capacitatea de a reproduce și transmite informații ereditare din generațiile anterioare către descendenți printr-un mecanism destul de simplu de autocopiare a macromoleculei de ADN a fiecărei celule vii. Fiecare, indiferent dacă organismul este format dintr-un număr mare de celule sau dacă vorbim despre ADN-ul care se găsește în celulele bacteriilor, aceste organisme simple unicelulare care nu sunt întotdeauna capabile să se adune chiar și într-o colonie mare. .
La fel ca toți reprezentanții vieții organice, informațiile ereditare (genetice) ale bacteriilor sunt stocate în ADN-ul lor. Ce este informația genetică? Ce structură stochează informații ereditare?
- Informația genetică este o secvență specifică de nucleotide. Nu există alt secret în nucleu. Prin copierea acestei secvențe, celula sintetizează o mare varietate de proteine. Ele rezolvă, de asemenea, toate celelalte probleme ale corpului, de la probleme de organizare până la alimentarea celulei cu materiale de construcție.
- Macromolecula de ADN este formată din patru baze nucleice (adenină, guanină, timină și citozină), unite într-o dublă spirală de zahăr dezoxiriboză și reziduuri de acid fosforic. Bazele nucleice codifică secvența de asamblare a proteinei, indiferent dacă există sau nu un nucleu format în celulă.
Acidul dezoxiribonucleic al bacteriilor are aceeași structură ca și moleculele care stochează informațiile ereditare ale tuturor celorlalte ființe vii de pe planetă. La fel ca toate celelalte celule organice, bacteriile formează cromozomi din ADN. Dar asta nu înseamnă că nu există alte diferențe.
Diferența fundamentală dintre o bacterie este că nu are un nucleu celular, informațiile ereditare ale unei bacterii nu sunt colectate în nucleul celulei, este pur și simplu o moleculă inel care este lipită de unul dintre pereții membranei citoplasmatice.
 Cu toate acestea, faptul că nu există un nucleu nu împiedică procesele active de replicare și traducere folosind acest păstrător de informații ereditare. Pentru a înțelege cum sunt transferate informațiile, trebuie să înțelegeți ce sunt cromozomii, genele și nucleul celular.
Cu toate acestea, faptul că nu există un nucleu nu împiedică procesele active de replicare și traducere folosind acest păstrător de informații ereditare. Pentru a înțelege cum sunt transferate informațiile, trebuie să înțelegeți ce sunt cromozomii, genele și nucleul celular.
- O genă este o secțiune a unei macromolecule pe care este scrisă o secvență de nucleotide care permite asamblarea unui anumit tip de proteină. Nu există alte informații în gene.
- Un cromozom este o combinație a unei catene de ADN cu proteine histone care o structurează și îi conferă o anumită formă înainte ca celula să înceapă să se divizeze. În faza în care nu are loc diviziunea, nu există cromozomi ca atare în celulă (sau în nucleu, dacă vorbim de eucariote nucleare).
- Nucleul celular este o structură celulară care conține informații ereditare structurate într-un cromozom atunci când celula se pregătește să se divizeze. Acesta inițiază procesul de divizare în sine. Este important să ne amintim că bacteriile nu au un nucleu celular.
Dacă într-o celulă eucariotă, în timpul diviziunii, se folosesc structuri separate, special formate pentru comoditatea diviziunii, atunci cum se reproduc bacteriile în condiții de haos neformat, aparent, în absența unui nucleu celular?
Acidul dezoxiribonucleic al unei celule bacteriene
Deși molecula de ADN bacteriană este descrisă ca o structură circulară destul de voluminoasă situată în centrul celulei, este de fapt o formațiune destul de compactă localizată în zone limitate ale citoplasmei.
 Datorită absenței unei membrane nucleare care să separe macromolecula bacteriană asamblată de alte structuri celulare, aparatul genetic al organismelor fără nucleu nu poate fi asociat cu aparatul genetic al eucariotelor, prin urmare aparatul genetic al procariotelor a fost numit nucleoid.
Datorită absenței unei membrane nucleare care să separe macromolecula bacteriană asamblată de alte structuri celulare, aparatul genetic al organismelor fără nucleu nu poate fi asociat cu aparatul genetic al eucariotelor, prin urmare aparatul genetic al procariotelor a fost numit nucleoid.
Trăsăturile caracteristice ale unui nucleoid:
- ADN care conține câteva mii de gene.
- Genele sunt aranjate liniar și se numesc cromozomi. Cromozomul unei bacterii este o colecție liniară a genelor sale.
- Macromolecula este, de asemenea, pliată de proteine asemănătoare histonelor eucariote.
Nucleoidul este atașat de membrana citoplasmatică în punctele în care începe și se termină replicarea (autocopiere).

S-a stabilit experimental că un nucleoid și un cromozom nu sunt același lucru. O creștere a numărului de cromozomi (gene liniare) este o dovadă că bacteriile se divid activ. Un nucleoid poate consta dintr-un cromozom sau mai multe copii ale acestuia. Astfel, în timpul perioadei de diviziune, Azotobacter se replic la 20-25 de cromozomi (copii nucleoide).
Procesul de copiere
În proiectele teoretice dezvoltate de microbiologi în acei ani în care era foarte dificil sau practic imposibil să se studieze procesele moleculare complexe în mod experimental, copierea acidului dezoxiribonucleic poate fi realizată în trei moduri:
- Conservator, în care helixul dublu părinte nu se desfășoară, iar helixul dublu fiică este format în întregime din material nou.
- Dispersiv, în care macromolecula părinte se descompune în fragmente, iar cele fiice se formează pe secvențele de nucleotide ale acestor fragmente ca pe șabloane.
- Semi-conservator. Conform acestui model, helixul dublu se desfășoară și fiecare șuviță a helixului servește ca șablon pentru ADN-ul fiicei. Se formează un așa-numit hibrid al unei macromolecule vechi și al unui lanț creat din noi componente.
Când în 1957 a fost găsită o modalitate de a monitoriza procesele care au loc în ADN-ul bacterian în timpul replicării acestuia, s-a constatat că acidul dezoxiribonucleic se replic într-un mod semi-conservator, adică prin derulare și utilizarea regiunilor de desfășurare ca șabloane pentru sinteza de noi macromolecule.

Procesul de replicare a ADN-ului bacterian în sine este foarte asemănător cu replicarea ADN-ului altor mecanisme organice. Se întâmplă după următoarea schemă:
- ADN-helicazele se desfășoară și sparg dubla helix prin mișcarea de-a lungul coloanei vertebrale zahăr-fosfat a acidului dezoxiribonucleic.
- Enzimele polimeraze catalizează adăugarea de baze nucleice complementare la fragmentele monocatenar de acid dezoxiribonucleic.
După replicare, toate părțile principale ale celulei sunt duplicate: organele, membrana citoplasmatică, peretele celular și celula bacteriană se împarte în două.
Probleme
Pe lângă interesul pur științific pentru studiul ADN-ului bacterian, mecanismul de replicare și transmitere a informațiilor ereditare de la o celulă la alta are și o importanță practică excepțională.
Este un fapt larg cunoscut că bacteriile se adaptează foarte repede atunci când sunt expuse la antibiotice și încep să producă anumite proteine anticorpi care blochează efectul distructiv al antibioticelor asupra celulei bacteriene. În generațiile ulterioare de bacterii, această rezistență la un grup specific de medicamente antibacteriene este menținută.
 Mai mult, prin transferul orizontal al genelor (nu prin divizare, ci prin simplul contact al unei bacterii cu alta), se transferă și o astfel de informație genetică, făcând un număr tot mai mare de specii bacteriene rezistente la antibiotice.
Mai mult, prin transferul orizontal al genelor (nu prin divizare, ci prin simplul contact al unei bacterii cu alta), se transferă și o astfel de informație genetică, făcând un număr tot mai mare de specii bacteriene rezistente la antibiotice.
Studiul acestor proprietăți ale bacteriilor, determinarea modului în care o genă străină este inclusă în structura generală a acidului dezoxiribonucleic, este ceea ce se ocupă microbiologia modernă.
GENETICA BACTERIILOR
Scopul prelegerii: familiarizarea studenților cu elementele de bază ale geneticii bacteriene și metodele de bază ale diagnosticului genetic.
Schema cursului
1. Caracteristicile organizării aparatului nuclear al bacteriilor.
2. Compoziția genomului bacterian.
3. Variabilitatea bacteriilor.
4. Recombinări în bacterii și caracteristicile acestora.
5. Diagnosticul genetic.
6. Clasificarea bacteriilor.
1. Caracteristici ale organizării morfologice a aparatului nuclear al bacteriilor:
- nu are membrană nucleară sau nucleol; se numește nucleoid;
- ADN-ul este purtătorul de informații genetice. Dacă la eucariote ADN-ul este liniar, atunci la majoritatea bacteriilor este circular, iar o catenă este fixată pe membrana citoplasmatică. Dacă desfășurați ADN-ul, lungimea acestuia va fi de sute de ori mai mare decât lungimea celulei. ADN-ul bacterian este supraînfăşurat.
- o celulă bacteriană conține un cromozom, adică bacteriile sunt organisme haploide.
2. Caracteristici biochimice.
- ADN-ul bacterian are aceeași compoziție ca ADN-ul eucariotic.
- În bacterii, ADN-ul poate conține baze minore, a căror prezență protejează ADN-ul de acțiunea propriilor endonucleaze.
- în genomul bacteriilor patogene există secțiuni de ADN care diferă de genomul principal prin compoziție perechi de baze nucleotide G-C. Aceste zone sunt responsabile pentru sinteza factorilor de patogenitate – insule de patogenitate.
- ADN-ul bacterian nu conține histone, iar rolul lor este jucat de poliamine. Genomul bacterian este reprezentat de structuri care sunt capabile de
replicare offline. Există două astfel de structuri: cromozomi, în care toate informațiile vitale sunt codificate (un cromozom bacterian conține până la 3 mii de gene diferite) și plasmide.
Plasmidele sunt ADN de natură circulară. Plasmidele dintr-o celulă pot fi în una din două stări alternative: libere sau integrate cu cromozomul.
Plasmidele codifică informații genetice suplimentare care nu sunt vitale pentru celulă, dar prezența acestor informații îi conferă anumite avantaje selective. Plasmidele includ:
gene structurale; -gene responsabile de propria replicare a plasmidei.
Unele plasmide au gene care asigură transmisibilitatea plasmidei - gene tra.
În funcție de atributul codificat, ei disting:
- Plasmidele R codifică rezistența bacteriană la medicamente;
- Plasmide F (sex) - determină capacitatea celulei de a fi donator de informații genetice;
- Plasmida Col - codifică sinteza bacteriocinelor;
- plasmide responsabile de sinteza factorilor de virulență(Ent-, Hly-)
și alte plasmide.
Genomul bacterian include elemente genetice mobile: elemente IS (secvențe de inserție), transpozoni și integroni. Ele se găsesc atât ca parte a cromozomului bacterian, cât și ca parte a plasmidelor. Replicarea lor este o parte integrantă a replicării cromozomilor și plasmidelor.
Elementele IS sunt secvențe de nucleotide scurte (2000). O trăsătură distinctivă a elementelor IS este prezența repetărilor inversate la capete, care sunt recunoscute de transpoză. Nu poartă gene structurale; sunt aceleași în bacterii din diferite specii, genuri și chiar se crede că sunt aceleași la procariote și eucariote. Elementele IS se pot deplasa atât de-a lungul unui cromozom, cât și între cromozomi. Acestea conțin 2 gene: prima codifică sinteza transpozazei; această enzimă asigură procesul de excludere a elementului IS din cromozom și integrarea acestuia în noul locus cromozom. A 2-a genă codifică sinteza unui represor, care reglează întregul proces de mișcare.
Transpozonii sunt segmente de ADN care au aceleași proprietăți ca și elementul IS, dar au gene structurale.
Integronii sunt elemente genetice mobile; conțin o genă care codifică rezistența la antibiotice. Integronii sunt un sistem de captare a elementelor mici de ADN numite casete genice prin recombinare specifică locului și expresia lor.
Semnificația elementelor mobile.
Deplasându-se de-a lungul ADN-ului unei celule sau între ADN-ul diferitelor celule, acestea provoacă:
- inactivarea genelor acelor secțiuni de ADN în care acestea, după ce s-au mutat, sunt integrate;
- deteriorarea materialului genetic;
- integrarea plasmidei în cromozom;
- distribuția unei gene într-o populație bacteriană.
Bacteriile, ca toate ființele vii, se caracterizează prin variabilitate. Variația la eucariote are loc pe verticală, la bacterii - atât pe verticală, cât și pe orizontală.
Există două tipuri de variabilitate: - fenotipică - genotipică.
Variabilitatea fenotipică se manifestă sub formă de modificări - aceasta este o schimbare a proprietăților celulei sub influența influențelor externe.
Modificările pot fi pe termen lung sau pe termen scurt. Modificările de modificare afectează marea majoritate a celulelor din populație.
Genotipic este mutația sau recombinarea. Mutațiile pot fi spontane sau induse.
Recombinarea este interacțiunea dintre doi genomi cu genotipuri diferite, ceea ce duce la formarea unui genom care combină genele donatorului și ale primitorului. În timpul procesului de recombinare, bacteriile sunt împărțite în mod convențional în celule donatoare, care transferă material genetic, și celule primitoare, care îl primesc. Recombinarea în bacterii este considerată a fi analogă cu reproducerea sexuală.
Caracteristicile recombinării în bacterii:
- nu există meioză. Nu se formează un zigot, ci un merazigot.
- întotdeauna dirijată de la donator la beneficiar.
- cantitatea de material genetic dintr-un recombinant este întotdeauna mai mare de unu. Recombinantul conține toată informația genetică a primitorului și a unei părți
informația genetică a donatorului.
La eucariote, există un singur mecanism de recombinare - meioza; În bacterii, există trei tipuri de recombinare:
1) Transformarea este schimbul de informații genetice folosind ADN pur.
2) Transducția este o metodă de transfer de informații genetice folosind fagi.
3) Conjugarea este o metodă de transfer a informațiilor genetice atunci când se formează punți citoplasmatice între două bacterii. În timpul conjugării
Aproape întregul genom poate trece în celula primitoare.
Metodele genetice sunt utilizate în scopuri practice atât pentru a detecta un microb în materialul studiat fără izolarea unei culturi pure, cât și pentru a determina poziția taxonomică a microbilor și pentru a efectua identificarea intraspecifică.
Secvențierea genomului– determinarea secvenței perechilor de nucleotide ADN.
Analiza restricțiilor- această metodă se bazează pe utilizarea enzimelor de restricție - acestea sunt endonucleaze care scindează molecula de ADN doar în anumite locuri. Dacă ADN-ul izolat dintr-un anumit material este tratat cu o anumită enzimă de restricție, aceasta va duce la formarea unui număr strict definit de fragmente de ADN de dimensiuni fixe.
Ribotiparea– vă permite să determinați tipul de bacterii. Secvența bazelor nucleotidice din operoni care codifică ARNr este caracterizată prin prezența atât a regiunilor conservate, care au o structură similară în diferite bacterii, cât și a secvențelor variabile, care sunt specifice genului și speciei și sunt markeri pentru identificarea genetică.
Hibridarea moleculară– utilizat în sistematica genetică. Această metodă ne permite să determinăm gradul de similitudine dintre diferitele ADN-uri.
PCR este utilizată pentru a detecta gene sau secvențe de nucleotide corespunzătoare care codifică fie o specie, fie o altă trăsătură.
Metoda PCR se bazează pe principiul complementarității și vă permite să creșteți (amplificați) cantitatea de probă de ADN testată. Această metodă are o sensibilitate extrem de mare și teoretic permite detectarea chiar și a unor molecule de ADN individuale în materialul studiat.
Tipuri de PCR:
- PCR în timp real; face posibilă determinarea numărului de fragmente de ADN prezente în material, adică efectuează analize cantitative;
- PCR multiplex – avantajul este că în amestecul de reacție pot fi introduse 2-4 sau mai multe perechi de primeri. Ele sunt caracteristice diverșilor agenți patogeni.
- PCR cu transcripție inversă – permite copierea ARN-ului agenților patogeni.
Cipurile ADN sunt cele mai recente tehnologii pentru metodele de hibridizare ale analizei genetice moleculare. Sunt purtători de oligonucleotide cunoscute (mai puțin de 20 de baze fiecare), complementare cu secțiuni ale genomului (sau genomului) aflate în studiu și care ocupă un loc specific (celulă). Dacă în proba de testat există fragmente din ADN-ul dorit, acestea hibridizează (se unesc după principiul complementarității) cu secvențele de nucleotide situate pe cip.
Clasificarea bacteriilor.
Unitatea taxonomică de bază a bacteriilor este specia. Pentru a desemna speciile din bacterii, se folosește nomenclatura dublă (binară).
O specie în bacterii este o colecție de bacterii înrudite care au proprietăți biologice similare și au o origine comună. În prezent, există 3 abordări pentru clasificarea bacteriilor:
1. Clasificare de rutină.
Ea formează baza ghidului pentru bacterii editat de Bergey.
2. Taxonomie numerică.
3. Sistematica genelor.
Concluzie: studenții sunt familiarizați cu elementele de bază ale geneticii bacteriene și cu metodele de bază de diagnosticare genetică.



")



