Жгутики - это длинные нитевидные образования на поверхности клетки, обеспечивающие ее активное пространственное перемещение. Несмотря на многообразие организмов, эти структуры внутри каждого надцарства (прокариот либо эукариот) характеризуются общей схемой строения.
Общая характеристика жгутиков
У доядерных организмов (бактерий и архей) жгутики - это основной способ передвижения. Среди эукариот эти локомоторные структуры в основном присутствуют у одноклеточных организмов, - простейших, но также характерны для гамет растений и животных. У некоторых многоклеточных беспозвоночных, - например, губок, - жгутики выполняют функцию перемещения жидкого субстрата относительно неподвижного клеточного пласта.
Морфологически жгутик состоит из закрепленного в толще клетки основания и длинной наружной нити, совершающей вращательные движения по спиральной траектории. Строение и механизм работы этих частей у прокариот и эукариот сильно отличаются, в связи с чем выделяют два соответствующих класса жгутиков.
Особенности жгутиков прокариот и эукариот
Наружную нить жгутика называют филаментом. У прокариот она состоит из белка флагеллина и пассивно движется за счет вращения базального мотора. Филамент ядерных клеток устроен значительно сложнее и благодаря взаимодействию белков тубулина и динеина способен изгибаться самостоятельно.
| у прокариот | у эукариот | |
| размеры органеллы (толщина, нм; длина, мкм) | 10-30 нм, 6-15 мкм | 200 нм, 100 мкм |
| белки жгутиковой нити | флагеллин | тубулин и динеин |
| мембрана вокруг филамента | отсутствует | присутствует |
| степень вращения | 360 ° | 180 ° |
| источник энергии | трансмембранный потенциал (у архей возможно АТФ) | АТФ |
| движение нити | пассивное | активное |
| субструктуры | филамент, крюк, базальное тельце | филамент, базальное тело (кинетосома) |
| строение филамента | сплошной (у архей) или полый белковый цилиндр | дублеты микротрубочек |
| структура базальной части | стержень, закрепленный в сложной системе колец или мембраноподобные органеллы (у архей) | триплеты микротрубочек |
Такое количество отличий свидетельствует об отсутствии гомологии между этими органоидами, то есть они не одинаковы по происхождению и строению, хоть и выполняют сходные функции.
Надцарство прокариот включает в себя царства архей и бактерий. Локомоторные структуры этих таксонов тоже не гомологичны друг другу, однако очень близки по строению. Жгутики архей изучены гораздо хуже.
Жгутики архей и бактерий
По способу перемещения подвижные бактерии подразделяются на плавающие и скользящие. Жгутики - это локомоторный орган плавающих микроорганизмов, позволяющий им развивать скорость от 20 до 200 мкм/сек.
Движение может быть спонтанным (если физико-химические характеристики среды одинаковы во всех направлениях) либо целенаправленным, когда бактерия стремится попасть в наиболее выгодные для нее условия. При адаптивном перемещении вращение базального мотора контролируется сенсорными системами.
По количеству и расположению жгутиков на клетке бактерии выделяют четыре морфологических типа микроорганизмов:
- монотрихи - имеют единственный жгутик;
- лофотрихи - характеризуются жгутиковым пучком на одном из клеточных полюсов;
- амфитрихи - имеют один или несколько жгутиков на обоих концах клетки;
- перитрихи - покрыты множеством жгутиков со всех сторон.

Тип жгутикования может быть как видовым признаком, так и результатом изменения условий культивирования или стадии жизненного цикла бактерии.
Жгутик архей во многом похож на бактериальный, однако имеет ряд отличий в ультраструктуре и механизме движения. Так, филамент у архей тоньше, построен из другого типа флагеллина, полый каналец в нити отсутствует. Длина крюка непостоянна, базальное тело имеет совсем другое строение и функционирует, скорее всего, на основе энергии АТФ. Археи движутся значительно медленнее бактерий.
Строение и функционирование бактериального жгутика
Жгутик бактерий образован тремя субструктурами: наружной нитью (филаментом), гибким сочленением (крюком) и базальным тельцем, заякоренным в клеточной оболочке. Синтез и сборку этих элементов кодируют около 50 fla-генов. За работу мотора отвечают mot-гены, а за адаптивные реакции - che-гены.
Филамент жгутика - это относительно жесткая белковая спираль, закрученная против часовой стрелки с образованием центрального полого канала диаметром до 3 нм. Такая конструкция способствует формированию спиральной траектории движения нити. По каналу филамента транспортируются молекулы флагеллина (FliC).
Крюк соединяет нить с базальным телом жгутика и состоит из двух типов белка: FlgE и FlgKl. Длина сочленения постоянна и составляет около 50 нм. Из-за изогнутой формы крюка при вращении мотора основание фибриллы описывает круг, благодаря чему возможно спиралевидное движение жгутика.
Базальное тельце закреплено в клеточной стенке и цитоплазматической мембране бактерий. Эта субструктура выполняет не только фиксирующую функцию, но и является мотором жгутика. Строение и локализация базального тельца зависят от типа клеточной стенки микроорганизма. У грамотрицательных бактерий оно состоит из двух внутренних (M и S) и двух внешних (P и L) колец, нанизанных на соединенный с крюком стержень. В состав базального тела также входит экспортная система, транспортирующая белковые элементы для сборки жгутика.
Структура базального мотора
В состав M-S-комплекса входят движущиеся структуры, называемые ротором, и переключатель направления вращения, который на более подробных схемах строения обозначают как С-кольцо. Вокруг ротора сосредоточены образованные белками MotAB ионные каналы - статоры. Мотор работает за счет энергии протонного (H +) или натриевого (Na +) градиента.

Расположение кольцевых субъединиц в клеточной оболочке соответствует следующей схеме:
- "М" - цитоплазматическая мембрана;
- "S" - периплазматическое пространство или клеточная стенка у Г + -бактерий;
- "P" - пептидогликановый слой;
- "L" - наружняя мембрана.
Внешние кольца P и L неподвижны и выполняют поддерживающую функцию. У грамположительных бактерий они отсутствуют.
Жгутик ядерной клетки
Эукариотический жгутик представляет собой цитоплазматический вырост клетки, состоящий из окруженной мембраной внешней части (ундулиподия) и погруженного в цитоплазму базального тела (кинетосомы).

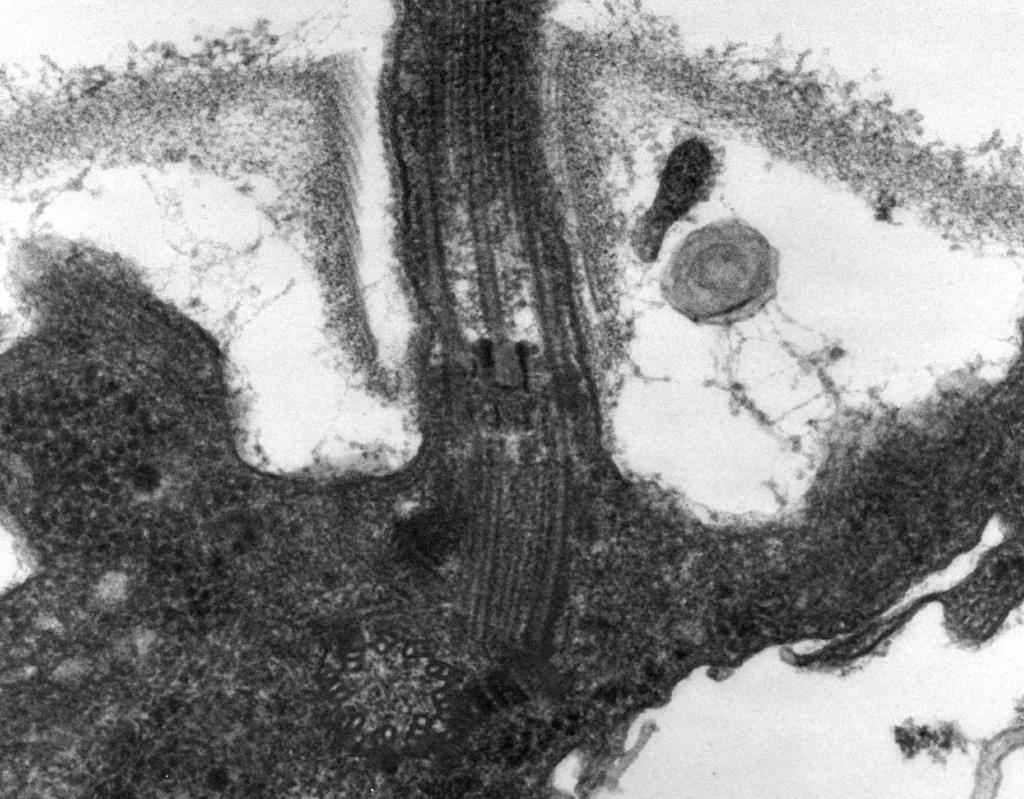
Структурной основой ундулиподия является аксонема, состоящая из системы связанных друг с другом белковых цилиндров - микротрубочек. Их расположение соответствует формуле 9×2+2, - то есть девять периферических дублетов и две одиночные трубочки в центре (синглеты).
Дублеты образованы спаренными цилиндрами А и В, построенными из субъединиц белка тубулина. От каждой А-трубочки в сторону соседней пары отходит динеиновые рукоятки, которые преобразуют энергию АТФ в механическое движение. Дублеты соединены с синглетами радиальными спицами, а друг с другом - нексиновыми связками. Пространство между структурными элементами ундулиподия заполнено цитоплазмой.

Структура кинетосомы представлена девятью триплетами микротрубочек (формула 9+0), которые заякоривают жгутик в эукариотической клетке. Синглеты в базальном теле отсутствуют.
Как прокариотические, так и эукариотические могут содержат структуры, известные как реснички и жгутики. Эти выросты на поверхности клеток помогают в их .
Особенности и функции
Реснички и жгутики являются выростами из некоторых клеток необходимые для клеточной локомоции (передвижения). Они также помогают перемещать вещества вокруг клеток и направлять их к нужным участкам.
Реснички и жгутики образуются из специализированных групп микротрубочек, называемых базальными телами.
Если выросты короткие и многочисленные, их называют ресничками. Если они длиннее и менее многочисленны (обычно только один или два), они называются жгутиками.
Строение
Обычно реснички и жгутики имеют сердцевину, состоящую из микротрубочек, соединенных с , расположенных по схеме 9+2. Кольцо из девяти микротрубочек имеет в своем центре две особые микротрубочки, которые сгибают реснички или жгутики. Этот тип организации встречается в устройстве большинства ресничек и жгутиков .
Где встречаются?
Как реснички, так и жгутики встречаются во многих типах клеток. Например, у спермы многих животных, водорослей и даже папоротников есть жгутики. Реснички можно найти в клетках таких тканей, как дыхательные пути и женский репродуктивный тракт.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
8. Основные формы бактерий
Шаровидные бактерии (кокки) имеют обычно форму шара, но могут быть немного овальной или бобовидной формы. Кокки могут располагаться поодиночке (микрококки); попарно (диплококки); в виде цепочек (стрептококки) или виноградных гроздьев (стафилококки), пакетом (сарцины). Стрептококки могут вызывать ангину и рожистое воспаление, стафилококки - различные воспалительные и гнойные процессы.
Палочковидные бактерии самые распространенные. Палочки могут быть одиночными, соединяться попарно (диплобактерии) или в цепочки (стрептобактерии). К палочковидным относятся кишечная палочка, возбудители сальмонеллеза, дизентерии, брюшного тифа, туберкулеза и др. Некоторые палочковидные бактерии обладают способностью при неблагоприятных условиях образовывать споры. Спорообразующие палочки называют бациллами. Бациллы, напоминающие по форме веретено, называют клостридиями.
Спорообразование представляет собой сложный процесс. Споры существенно отличаются от обычной бактериальной клетки. Они имеют плотную оболочку и очень малое количество воды, им не требуются питательные вещества, а размножение полностью прекращается. Споры способны длительно выдерживать высушивание, высокие и низкие температуры и могут находиться в жизнеспособном состоянии десятки и сотни лет (споры сибирской язвы, ботулизма, столбняка и др.). Попав в благоприятную среду, споры прорастают, т. е. превращаются в обычную вегетативную размножающуюся форму.
Извитые бактерии могут быть в виде запятой - вибрионы, с несколькими завитками - спириллы, в виде тонкой извитой палочки - спирохеты. К вибрионам относится возбудитель холеры, а возбудитель сифилиса - спирохета.
9. Особенности морфологии риккетсий и хламидий
Риккетсии представляют собой мелкие грамотрицательные микроорганизмы, характеризующиеся выраженным полиморфизмом-образуют кокковидные, палочковидные и нитевидные формы (рис. 22). Размеры риккетсии варьируют от 0,5 до 3-4 мкм, длина нитевидных форм достигает 10-40 мкм. Спор и капсул не образуют, окрашиваются по Здродовскому в красный цвет.
Хламидии имеют шаровидную, овоидную или палочковидную форму. Их размеры колеблются в пределах 0,2-1,5 мкм. Морфология и размеры хламидии зависят от стадии их внутриклеточного цикла развития, для которого характерно превращение небольшого шаровидного элементарного образования в крупное инициальное тельце с бинарным делением. Перед делением частицы хламидии обволакиваются образованием, напоминающим бактериальную капсулу. Хламидии окрашиваются"* по Романовскому-Гимзе, грамотрицательны, хорошо видны в прижизненных препаратах при фазово-контрастной микроскопии.
10. Строение и биология микоплазм.
К классу Mollicutes относится только один порядок Mycoplasmatales. Представители этого порядка - микоплазмы-
отличаются от бактерий отсутствием клеточной стенки. Вместо нее они содержат трехслойную липопротеидную цитоплазматическую мембрану. Размеры микоплазм колеблются в пределах 125-250 мкм. Они имеют форму круглых, овальных или нитевидных образований, грамотрицательны
Микоплазмы размножаются бинарным делением, подобно большинству бактерий, особенно после образования мелких кокковидных образований (элементарные тельца, ЭТ) в нитевидных структурах.
Микоплазмы способны к почкованию и сегментации. Минимальной репродуцирующейся единицей считают ЭТ (0,7-0,2 мкм). Основной компонент клеточной мембраны - холестерин. Микоплазмы не способны к образованию холестерина и утилизируют его из тканей или питательных сред, дополненных их внесением. По Граму окрашиваются отрицательно, но лучшие результаты даёт окраска по Романовскому-Гимзе. Микоплазмы требовательны к условиям культивирования: в питательные среды необходимо вносить нативную сыворотку, холестерин, нуклеиновые кислоты, углеводы, витамины и различные соли. На плотных средах образуют характерные мелкие полупрозрачные колонии с приподнятым зернистым центром, придающим им вид «яичницы-глазуньи». На средах с кровью некоторые виды микоплазм дают а- и бета-гемолиз. В полужидких средах микоплазмы растут по линии укола, формируя дисперсные, крош-коватые колонии. В жидких средах приводят к незначительному помутнению или опалесценции; некоторые штаммы способны образовывать тончайшую жирную плёнку. У человека выделяют представителей родов Mycoplasma, Ureaplasma и Acholeplasma, включающих патогенные и сапрофитические виды
От переднего полюса тела берут начало жгутики (1, 2, 4, 8 и более - до нескольких тысяч). Если их много, они могут покрывать все тело простейшего (например, в отряде Hypermastigina и отряде Opalinina), напоминая тем самым инфузорий . Длина жгутиков варьирует в широких пределах - от немногих до нескольких десятков микрометров. Если жгутов два, то нередко один выполняет локомоторную функцию, а второй неподвижно тянется вдоль тела и выполняет функцию руля. У некоторых жгутиконосцев (род Trichomonas, род Trypanosoma) жгутик проходит вдоль тела ( рис. 19) и соединяется с последним при помощи тонкой цитоплазматической перепонки. Таким образом формируется ундулирующая мембрана, которая волнообразными колебаниями вызывает поступательное движение простейшего.
В деталях механизм работы жгутиков различен, но в основе это винтообразное движение. Простейшее как бы "ввинчивается" в окружающую среду. Жгутик совершает от 10 до 40 об/с.
Ультраструктура жгутиков очень сложна и обнаруживает поразительное постоянство во всем животном и растительном мире. Все жгутики и реснички животных и растений построены по единому плану (за единичными отклонениями) ( табл. I).
Каждый жгутик слагается из двух отделов. Большую часть его составляет свободный участок, отходящий от поверхности клетки наружу и являющийся собственно локомоторным. Второй отдел жгутика - базальное тело (кинетосома) - меньшая по размерам часть, погруженная в толщу эктоплазмы. Снаружи жгутик покрыт трехслойной мембраной, представляющей непосредственное продолжение наружной мембраны клетки.
Внутри жгутика строго закономерно располагаются 11 фибрилл. Вдоль оси жгута проходят 2 центральные фибриллы ( рис. 20), берущие начало от аксиальной гранулы. Диаметр каждой из них составляет около 25 нм, а центры их расположены на расстоянии 30 нм. По периферии под оболочкой находятся еще 9 фибрилл, причем каждая слагается из двух тесно спаянных трубочек. Локомоторная деятельность жгутика определяется периферическими фибриллами, тогда как центральные играют опорную функцию и может быть представляют собой субстрат, по которому распространяются волны возбуждения, вызывающие движение жгутика.
Базальное тело (кинетосома) , находится в эктоплазме. Оно имеет вид цилиндрического тельца, окруженного мембраной, под которой по периферии располагаются 9 фибрилл, являющихся непосредственным продолжением периферических фибрилл самого жгута. Здесь они, правда, становятся тройными ( рис. 20 , табл. II). Иногда основание жгутика продолжается в глубину цитоплазмы за пределы кинетосомы, образуя корневую нить (ризопласт), которая может либо свободно заканчиваться в цитоплазме, либо прикрепляться к оболочке ядра.
У некоторых жгутиконосцев вблизи кинетосомы располагается парабазальное тело . Форма его может быть разнообразной. Иногда это яйцевидное или колбасовидное образование, иногда же оно приобретает довольно сложную конфигурацию и состоит из множества отдельных долек (
Развитие микробиологии принесло в последние десятилетия множество открытий. И одно из них - это особенности движения жгутиковых бактерий. Устройство двигателей этих древнейших организмов оказалось очень сложным и по принципу своей работы очень отличающимся от жгутиков наших ближайших эукариотических родственников простейших. Вокруг двигателя жгутиковой бактерии разгорелись самые жаркие споры между креационистами и эволюционистами. О бактериях, их жгутиковых моторах и о многом другом - данная статья.
Общая биология
Для начала вспомним, что это за организмы и какое место они занимают в системе органического мира на нашей планете. Домен Bacteria объединяет огромное количество одноклеточных прокариотических (без оформленного ядра) организмов.
Эти живые клетки появились на арене жизни почти 4 миллиарда лет назад и были первыми поселенцами планеты. Они могут быть самой разной формы (кокки, палочки, вибрионы, спирохеты), но большинство из них - жгутиковые.
Где живут бактерии? Везде. На планете их обитает более 5×10 30 . В 1 грамме почвы их насчитывается порядка 40 миллионов, в нашем организме обитает до 39 триллионов. Их можно встретить на дне Марианской впадины, в горячих «черных курильщиках» на дне океанов, во льдах Антарктиды, а на ваших руках в данный момент находится до 10 миллионов бактерий.
Значение неоспоримо
Несмотря на микроскопические размеры (0,5-5 мкм) их общая биомасса на Земле больше, чем биомасса животных и растений, вместе взятых. Их роль в круговороте веществ незаменима, а их свойства консументов (разрушителей органики) не позволяют планете покрыться горами трупов.
Ну и не стоит забывать о патогенах: возбудителями чумы, оспы, сифилиса, туберкулеза и множества других инфекционных заболеваний тоже являются бактерии.
Бактерии нашли применение и в хозяйственной деятельности человека. Начиная от пищевой промышленности (кисломолочные продукты, сыры, квашеные овощи, алкогольные напитки), «зеленой» экономике (биотопливо и биогаз) до методов клеточной инженерии и производства лекарственных препаратов (вакцины, сыворотки, гормоны, витамины).

Общая морфология
Как уже говорилось, у этих одноклеточных представителей жизни нет ядра, их наследственный материал (молекулы ДНК в виде кольца) расположены в определенном участке цитоплазмы (нуклеоид). Клетка их имеет плазматическую мембрану и плотную капсулу, образованную пептидогликаном муреином. Из клеточных органелл у бактерий есть митохондрии, могут быть хлоропласты и другие структуры с различными функциями.
Большинство бактерий - жгутиковые. Плотная капсула на поверхности клетки не позволяет им передвигаться при помощи изменения самой клетки, как это делают амебы. Их жгутики - это плотные белковые образования различной длины и диаметром около 20 нм. Одни бактерии имеют единственный жгутик (монотрихи), у других их два (амфитрихи). Иногда жгутики расположены пучками (лофотрихи) или же покрывают всю поверхность клетки (перитрихи).
Многие из них живут в виде одиночных клеток, но некоторые образуют скопления (пары, цепочки, филаменты, гифы).

Особенности движения
Жгутиковые бактерии могут двигаться по-разному. Одни движутся только вперед, а направление меняют путем кувыркания. Некоторые способны к подергиваниям, другие передвигаются путем скольжения.
Жгутики бактерий выполняют функции не только клеточного «весла», но могут быть и «абордажным» орудием.
Совсем до недавнего времени считали, что жгутик бактерии виляет как хвост у змеи. Последние исследования показали, что жгутик бактерий устроен куда сложнее. Он работает как турбина. Присоединенный к приводу, он вращается в одном направлении. Привод, или жгутиковый мотор бактерии, - это сложная молекулярная структура, которая работает как мышца. С тем отличием, что мышца после совершения сжатия должна расслабляться, а мотор бактерии работает постоянно.

Наномеханизм работы жгутика
Не углубляясь в биохимию движения, отметим, что в создании привода жгутика учувствует до 240 белков, которые делятся на 50 молекулярных составляющих с определенной функцией в системе.
В этой двигательной системе бактерии есть ротор, который движется, и статор, который обеспечивает это движение. Есть приводной вал, втулка, сцепление, тормоза и ускорители
Этот миниатюрный двигатель позволяет бактерии совершать перемещения на расстояния в 35 раз большие, чем ее собственный размер всего лишь за 1 секунду. При этом на работу самого жгутика, совершающего 60 тысяч оборотов в минуту, организм тратит всего 0,1 % всей энергии, которую расходует клетка.
Удивительно еще и то, что все запчасти своего двигательного механизма бактерия может заменять и ремонтировать «на ходу». Просто представьте, что вы летите в самолете. А техники меняют лопасти работающего мотора.
Жгутиковая бактерия против Дарвина
Двигатель, способный работать со скоростью до 60 000 оборотов в минуту, самочинящийся и использующий в виде топлива всего лишь углеводы (сахара), имеющий устройство сродни электрическому двигателю - могло ли такое устройство возникнуть в процессе эволюции?
Именно этот вопрос задал себе в 1988 году доктор биологических наук Майкл Бихи. Он ввел в биологию понятие несократимой системы - системы, в которой все ее части одновременно необходимы для обеспечения ее работы, а удаление хоть одной части приводит к полному нарушению ее функционирования.
С позиции эволюции Дарвина, все структурные изменения в организме происходят постепенно и отбираются естественным отбором только успешные.
Выводы М. Бихи, изложенные в книге «Черный ящик Дарвина» (1996): двигатель жгутиковой бактерии - неделимая система из более чем 40 частей, и отсутствие хоть одной приведет к полной нефункциональности системы, а значит, эта система не могла произойти путем естественного отбора.

Бальзам для креационистов
Теория сотворения в изложении ученого и профессора биологии, декана факультета биологических наук в Лихайском университете Бетлехема (США) М. Бихи сразу привлекла внимание служителей церкви и сторонников теории божественного происхождения жизни.
В 2005 году в США даже состоялся судебный процесс, где Бихи выступал свидетелем со стороны сторонников теории «разумного дизайна», на котором рассматривался вопрос введения в школах Дувра изучения креационизма на курсе «О пандах и людях». Процесс был проигран, преподавание такого предмета признали противоречащим действующей конституции.
Но споры креационистов и эволюционистов продолжаются и сегодня.







