ពួកវាបង្កើតបាន 20-30% នៃម៉ាសកោសិកា។ ទាំងនេះរួមមាន biopolymers - ប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត ខ្លាញ់ ATP ជាដើម។
ប្រភេទផ្សេងគ្នានៃកោសិកាមានបរិមាណផ្សេងគ្នានៃសមាសធាតុសរីរាង្គ។ កាបូអ៊ីដ្រាតស្មុគស្មាញ គ្របដណ្តប់លើកោសិការុក្ខជាតិ ប្រូតេអ៊ីន និងខ្លាញ់លើសនៅក្នុងសត្វ។ ទោះជាយ៉ាងណាក៏ដោយ ក្រុមនីមួយៗនៃសារធាតុសរីរាង្គនៅក្នុងប្រភេទកោសិកាណាមួយអនុវត្តមុខងារដូចខាងក្រោមៈ ការផ្តល់ថាមពល ធ្វើជាសម្ភារៈសំណង់ ផ្ទុកព័ត៌មាន។ល។
កំប្រុក។ក្នុងចំណោមសារធាតុសរីរាង្គនៃកោសិកា ប្រូតេអ៊ីនកាន់កាប់កន្លែងដំបូងក្នុងន័យបរិមាណ និងតម្លៃ។ នៅក្នុងសត្វពួកគេមាន 50% នៃម៉ាស់ស្ងួតនៃកោសិកា។
នៅក្នុងខ្លួនមនុស្សមានម៉ូលេគុលប្រូតេអ៊ីនជាច្រើនប្រភេទដែលខុសគ្នាពីគ្នាទៅវិញទៅមក និងពីប្រូតេអ៊ីននៃសារពាង្គកាយផ្សេងទៀត។
| |
ចំណង Peptide៖
នៅពេលបញ្ចូលគ្នា ម៉ូលេគុលបង្កើតបាន: dipeptide, tripeptide ឬ polypeptide ។ វាគឺជាសមាសធាតុនៃអាស៊ីតអាមីណូ 20 ឬច្រើនជាងនេះ។ លំដាប់នៃការផ្លាស់ប្តូរអាស៊ីតអាមីណូនៅក្នុងម៉ូលេគុលគឺមានភាពចម្រុះបំផុត។ នេះអនុញ្ញាតឱ្យមានអត្ថិភាពនៃវ៉ារ្យ៉ង់ដែលខុសគ្នានៅក្នុងតម្រូវការនិងលក្ខណៈសម្បត្តិនៃម៉ូលេគុលប្រូតេអ៊ីន។
លំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងម៉ូលេគុលត្រូវបានគេហៅថារចនាសម្ព័ន្ធ។
បឋម - លីនេអ៊ែរ។
អនុវិទ្យាល័យ - វង់។
កម្រិតទីបី - globules ។
Quaternary - សមាគមនៃ globules (អេម៉ូក្លូប៊ីន) ។
ការបាត់បង់រចនាសម្ព័ន្ធដោយម៉ូលេគុលត្រូវបានគេហៅថា denaturation ។ វាបណ្តាលមកពីការផ្លាស់ប្តូរសីតុណ្ហភាព pH វិទ្យុសកម្ម។ ជាមួយនឹងផលប៉ះពាល់បន្តិចបន្តួចម៉ូលេគុលអាចស្ដារលក្ខណៈសម្បត្តិរបស់វា។ វាត្រូវបានគេប្រើក្នុងថ្នាំ (ថ្នាំអង់ទីប៊ីយោទិច) ។
មុខងាររបស់ប្រូតេអ៊ីននៅក្នុងកោសិកាមានភាពចម្រុះ។ សំខាន់បំផុតគឺការសាងសង់។ ប្រូតេអ៊ីនត្រូវបានចូលរួមនៅក្នុងការបង្កើតភ្នាសកោសិកាទាំងអស់នៅក្នុងសរីរាង្គ។ មុខងារកាតាលីករមានសារៈសំខាន់ខ្លាំងណាស់ - អង់ស៊ីមទាំងអស់គឺជាប្រូតេអ៊ីន។ មុខងារម៉ូទ័រត្រូវបានផ្តល់ដោយប្រូតេអ៊ីន contractile ។ ការដឹកជញ្ជូន - មាននៅក្នុងការភ្ជាប់ធាតុគីមីនិងផ្ទេរពួកវាទៅជាលិកា។ មុខងារការពារត្រូវបានផ្តល់ដោយប្រូតេអ៊ីនពិសេស - អង្គបដិប្រាណដែលបង្កើតឡើងនៅក្នុង leukocytes ។ ប្រូតេអ៊ីនបម្រើជាប្រភពថាមពល - ជាមួយនឹងការបំបែកពេញលេញនៃប្រូតេអ៊ីន 1 ក្រាម 11.6 kJ ត្រូវបានបញ្ចេញ។
កាបូអ៊ីដ្រាត។ទាំងនេះគឺជាសមាសធាតុកាបូន អ៊ីដ្រូសែន និងអុកស៊ីសែន។ តំណាងដោយជាតិស្ករ។ កោសិកាមានរហូតដល់ 5% ។ អ្នកមានបំផុត - កោសិការុក្ខជាតិ - រហូតដល់ 90% នៃម៉ាស (ដំឡូងបារាំង) ។ ពួកគេត្រូវបានបែងចែកជាសាមញ្ញនិងស្មុគស្មាញ។ សាមញ្ញ - monosugar (គ្លុយកូស) C 6 H 12 O 6, ស្ករទំពាំងបាយជូ, fructose ។ Disahara - (sucrose) C] 2 H 22 O 11 beet និងស្ករអំពៅ។ Polysugar (សែលុយឡូស ម្សៅ) (C 6 H 10 O 5) n.
កាបូអ៊ីដ្រាតអនុវត្តមុខងារជាចម្បងនៃអគារ និងថាមពល។ នៅពេលដែលកាបូអ៊ីដ្រាត 1 ក្រាមត្រូវបានកត់សុី 17.6 kJ ត្រូវបានបញ្ចេញ។ ម្សៅ និង glycogen បម្រើជាទុនបម្រុងថាមពលរបស់កោសិកា។
លីពីត។ទាំងនេះគឺជាសារធាតុខ្លាញ់ និងសារធាតុខ្លាញ់នៅក្នុងកោសិកា។ ពួកវាជា esters នៃ glycerol និងអាស៊ីតឆ្អែត និង unsaturated ទម្ងន់ម៉ូលេគុលខ្ពស់។ ពួកវាអាចរឹងនិងរាវ - ប្រេង។ រុក្ខជាតិមាននៅក្នុងគ្រាប់ពូជពី 5-15% នៃសារធាតុស្ងួត។
មុខងារសំខាន់គឺថាមពល - នៅពេលដែល 1 ក្រាមនៃជាតិខ្លាញ់ត្រូវបានបំបែក 38.9 kJ ត្រូវបានបញ្ចេញ។ ខ្លាញ់គឺជាឃ្លាំងផ្ទុកសារធាតុចិញ្ចឹម។ ខ្លាញ់អនុវត្តមុខងារអគារគឺជាអ៊ីសូឡង់កំដៅដ៏ល្អ។
អាស៊ីតនុយក្លេអ៊ីក។ទាំងនេះគឺជាសមាសធាតុសរីរាង្គដ៏ស្មុគស្មាញ។ ពួកវាមាន C, H 2, O 2, N 2, P. ដែលមាននៅក្នុង nuclei និង cytoplasm ។
 |
ក) DNA គឺជាប៉ូលីណុក្លេអូទីតជីវសាស្រ្តដែលមានខ្សែសង្វាក់ពីរនៃនុយក្លេអូទីត។ នុយក្លេអូទីត - មានមូលដ្ឋានអាសូតចំនួន 4: 2 purines - Adenine និង Valine, 2 pyrimedins Cytosine និង Guanine ក៏ដូចជាស្ករ - deoxyribose និងសំណល់អាស៊ីតផូស្វ័រ។
នៅក្នុងខ្សែសង្វាក់នីមួយៗ នុយក្លេអូទីតត្រូវបានតភ្ជាប់ដោយចំណង covalent ។ ខ្សែសង្វាក់នៃ nucleotides បង្កើតជា helices ។ helix នៃ DNA ដែលផ្ទុកដោយប្រូតេអ៊ីនបង្កើតជារចនាសម្ព័ន្ធ - ក្រូម៉ូសូម។
ខ) RNA គឺជាវត្ថុធាតុ polymer ដែលជាម៉ូណូមឺរដែលជានុយក្លេអូទីតនៅជិត DNA មូលដ្ឋានអាសូត - A, G, C. ជំនួសឱ្យ thymine មាន Uration ។ កាបូអ៊ីដ្រាតនៃ RNA គឺ ribose មានសំណល់នៃអាស៊ីតផូស្វ័រ។

RNAs ពីរខ្សែគឺជាអ្នកបញ្ជូនព័ត៌មានហ្សែន។ ខ្សែតែមួយ - ផ្ទុកព័ត៌មានអំពីលំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីន។ មាន RNA ខ្សែតែមួយជាច្រើន៖
Ribosomal - នុយក្លេអូទីត 3-5 ពាន់;
ព័ត៌មាន - 300-30000 nucleotides;
ការដឹកជញ្ជូន - 76-85 nucleotides ។
ការសំយោគប្រូតេអ៊ីនត្រូវបានអនុវត្តនៅលើ ribosomes ដោយមានការចូលរួមពី RNA គ្រប់ប្រភេទ។
សំណួរសាកល្បង
1. កោសិកា - សារពាង្គកាយមួយឬផ្នែករបស់វា?
2. សមាសភាពបឋមនៃកោសិកា។
3. ទឹក និងសារធាតុរ៉ែ។
4. សារធាតុសរីរាង្គនៃកោសិកា។
សត្វ ផ្សិត និងបាក់តេរី
| សញ្ញា | បាក់តេរី | សត្វ | ផ្សិត | រុក្ខជាតិ |
| វិធីសាស្រ្តចិញ្ចឹម | heterotrophic ឬ autotrophic | Heterotrophic | Heterotrophic | អូតូត្រូហ្វីក |
| អង្គការ តំណពូជ ព័ត៌មាន | prokaryotes | eukaryotes | eukaryotes | eukaryotes |
| ការធ្វើមូលដ្ឋានីយកម្ម DNA | នុយក្លេអ៊ីត, ប្លាស្មា | ស្នូល, មីតូខនឌ្រី | ស្នូល, មីតូខនឌ្រី | នុយក្លេអ៊ែរ, មីតូខនឌ្រី, ផ្លាស្ទីត |
| ភ្នាសប្លាស្មា | មាន | មាន | មាន | មាន |
| ជញ្ជាំងកោសិកា | Mureinovaya | - | Chitinous | សែលុយឡូស |
| ស៊ីតូប្លាស្មា | មាន | មាន | មាន | មាន |
| សរីរាង្គ | រីបូសូម | Membrane និង non-membrane រួមទាំងមជ្ឈមណ្ឌលកោសិកា | ភ្នាស និងមិនមែនភ្នាស | Membrane និង non-membrane រួមទាំង plastids |
| សរីរាង្គនៃចលនា | Flagella និង villi | Flagella និង cilia | Flagella និង cilia | Flagella និង cilia |
| Vacuoles | កម្រណាស់។ | contractile, រំលាយអាហារ | ពេលខ្លះ | vacuole កណ្តាលជាមួយបឹងទន្លេសាប |
| ការរួមបញ្ចូល | វ៉ុលយូទីន | គ្លីកូហ្សែន | គ្លីកូហ្សែន | ម្សៅ |
ភាពខុសគ្នានៃរចនាសម្ព័ន្ធកោសិកានៃអ្នកតំណាងនៃនគរផ្សេងៗគ្នានៃសត្វព្រៃត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ២.៣.
អង្ករ។ ២.៣. រចនាសម្ព័ន្ធកោសិកានៃបាក់តេរី (A) សត្វ (B) ផ្សិត (C) និងរុក្ខជាតិ (D)
២.៣. អង្គការគីមីនៃកោសិកា។ ទំនាក់ទំនងនៃរចនាសម្ព័ន្ធ និងមុខងារនៃសារធាតុអសរីរាង្គ និងសរីរាង្គ (ប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត lipid ATP) ដែលបង្កើតជាកោសិកា។ យុត្តិកម្មនៃទំនាក់ទំនងនៃសារពាង្គកាយដោយផ្អែកលើការវិភាគនៃសមាសធាតុគីមីនៃកោសិការបស់ពួកគេ។
សមាសធាតុគីមីនៃកោសិកា។
នៅក្នុងសមាសភាពនៃសារពាង្គកាយមានជីវិត ភាគច្រើននៃធាតុគីមីនៃតារាងតាមកាលកំណត់នៃធាតុនៃ D. I. Mendeleev ដែលបានរកឃើញរហូតមកដល់បច្ចុប្បន្នត្រូវបានរកឃើញ។ ម៉្យាងវិញទៀត ពួកវាមិនមានធាតុតែមួយដែលមិនមាននៅក្នុងធម្មជាតិគ្មានជីវិតទេ ហើយម្យ៉ាងវិញទៀតការប្រមូលផ្តុំរបស់ពួកគេនៅក្នុងរូបកាយនៃធម្មជាតិគ្មានជីវិត និងសារពាង្គកាយមានជីវិតមានភាពខុសគ្នាខ្លាំង (តារាង 2.2)។
ធាតុគីមីទាំងនេះបង្កើតជាសារធាតុអសរីរាង្គ និងសារធាតុសរីរាង្គ។ ទោះបីជាការពិតដែលថាសារធាតុអសរីរាង្គមានឥទ្ធិពលលើសារពាង្គកាយមានជីវិត (រូបភាព 2.4) វាគឺជាសារធាតុសរីរាង្គដែលកំណត់ភាពប្លែកនៃសមាសធាតុគីមីរបស់វា និងបាតុភូតនៃជីវិតជាទូទៅ ដោយសារពួកវាត្រូវបានសំយោគជាចម្បងដោយសារពាង្គកាយនៅក្នុងដំណើរការនៃជីវិត និងការលេង។ តួនាទីសំខាន់ក្នុងប្រតិកម្ម។
វិទ្យាសាស្រ្តទាក់ទងនឹងការសិក្សាអំពីសមាសធាតុគីមីនៃសារពាង្គកាយ និងប្រតិកម្មគីមីដែលកើតឡើងនៅក្នុងពួកវា។ ជីវគីមី។
វាគួរតែត្រូវបានកត់សម្គាល់ថាមាតិកានៃសារធាតុគីមីនៅក្នុងកោសិកានិងជាលិកាផ្សេងគ្នាអាចប្រែប្រួលយ៉ាងខ្លាំង។ ឧទាហរណ៍ ប្រសិនបើប្រូតេអ៊ីននាំមុខក្នុងចំណោមសមាសធាតុសរីរាង្គនៅក្នុងកោសិកាសត្វ នោះកាបូអ៊ីដ្រាតលើសនៅក្នុងកោសិការុក្ខជាតិ។
តារាង 2.2
ខ្លឹមសារនៃធាតុគីមីមួយចំនួននៅក្នុងធម្មជាតិគ្មានជីវិត និងសារពាង្គកាយមានជីវិត,%
| ធាតុគីមី | សំបកផែនដី | ទឹកសមុទ្រ | សារពាង្គកាយមានជីវិត |
| អំពី | 49,2 | 85,8 | 65-75 |
| ពី | 0,4 | 0,0035 | 15-18 |
| ហ | 1,0 | 10,67 | 8-10 |
| ន | 0,04 | 0,37 | 1,5-3,0 |
| រ | 0,1 | 0,003 | 0,20-1,0 |
| ស | 0,15 | 0,09 | 0,15-0,2 |
| TO | 2,35 | 0,04 | 0,15-0,4 |
| ស | 3,25 | 0,05 | 0,04-2,0 |
| គ១ | 0,2 | 0,06 | 0,05-0,1 |
| មីលីក្រាម | 2,35 | 0,14 | 0,02-0,03 |
| ណា | 2,4 | 1.14 | 0,02-0,03 |
| ហ្វេ | 4,2 | 0,00015 | 0,01-0,015 |
| Zn | | 0,00015 | 0,0003 |
| គ | | | 0,0002 |
| ខ្ញុំ | | 0,000015 | 0,0001 |
| ច | 0,1 | 2,07 | 0,0001 |
ម៉ាក្រូ និងមីក្រូធាតុ
ធាតុគីមីប្រហែល 80 ត្រូវបានរកឃើញនៅក្នុងសារពាង្គកាយមានជីវិត ប៉ុន្តែមានតែ 27 ធាតុទាំងនេះប៉ុណ្ណោះដែលមានមុខងាររបស់វានៅក្នុងកោសិកា និងសារពាង្គកាយ។ ធាតុដែលនៅសេសសល់គឺមានវត្តមាននៅក្នុងបរិមាណដាន ហើយហាក់ដូចជាត្រូវបានបញ្ចូលតាមរយៈអាហារ ទឹក និងខ្យល់។ មាតិកានៃធាតុគីមីនៅក្នុងរាងកាយប្រែប្រួលយ៉ាងខ្លាំង (សូមមើលតារាង 2.2) ។ អាស្រ័យលើកំហាប់ពួកវាត្រូវបានបែងចែកទៅជា macronutrients និង microelements ។
ការផ្តោតអារម្មណ៍នីមួយៗ macronutrientsនៅក្នុងរាងកាយលើសពី 0.01% ហើយមាតិកាសរុបរបស់ពួកគេគឺ 99% ។ Macronutrients រួមមាន អុកស៊ីហ្សែន កាបូន អ៊ីដ្រូសែន អាសូត ផូស្វ័រ ស្ពាន់ធ័រ ប៉ូតាស្យូម កាល់ស្យូម សូដ្យូម ក្លរីន ម៉ាញេស្យូម និងជាតិដែក។ ធាតុទាំងបួនដំបូង (អុកស៊ីហ្សែន កាបូន អ៊ីដ្រូសែន និងអាសូត) ត្រូវបានគេហៅថាផងដែរ។ សរីរាង្គ,ដោយសារតែពួកវាគឺជាផ្នែកមួយនៃសមាសធាតុសរីរាង្គសំខាន់ៗ។ ផូស្វ័រ និងស្ពាន់ធ័រ ក៏ជាសមាសធាតុនៃសារធាតុសរីរាង្គមួយចំនួនផងដែរ ដូចជាប្រូតេអ៊ីន និងអាស៊ីតនុយក្លេអ៊ីក។ ផូស្វ័រមានសារៈសំខាន់សម្រាប់ការបង្កើតឆ្អឹង និងធ្មេញ។
បើគ្មានសារធាតុ macronutrients ដែលនៅសល់ មុខងារធម្មតារបស់រាងកាយគឺមិនអាចទៅរួចទេ។ ដូច្នេះប៉ូតាស្យូមសូដ្យូមនិងក្លរីនត្រូវបានចូលរួមនៅក្នុងដំណើរការនៃការរំភើបនៃកោសិកា។ ប៉ូតាស្យូមក៏ត្រូវការសម្រាប់អង់ស៊ីមជាច្រើនដើម្បីដំណើរការ និងរក្សាទឹកនៅក្នុងកោសិកា។ កាល់ស្យូមត្រូវបានរកឃើញនៅក្នុងជញ្ជាំងកោសិកានៃរុក្ខជាតិ ឆ្អឹង ធ្មេញ និងសំបក mollusk ហើយត្រូវបានទាមទារសម្រាប់ការកន្ត្រាក់សាច់ដុំ និងចលនាខាងក្នុង។ ម៉ាញ៉េស្យូមគឺជាសមាសធាតុនៃក្លរ៉ូហ្វីល - សារធាតុពណ៌ដែលធានាលំហូរនៃការធ្វើរស្មីសំយោគ។ វាក៏ចូលរួមក្នុងការសំយោគប្រូតេអ៊ីនផងដែរ។ ជាតិដែក បន្ថែមពីលើការជាផ្នែកមួយនៃអេម៉ូក្លូប៊ីន ដែលផ្ទុកអុកស៊ីហ្សែនក្នុងឈាម គឺចាំបាច់សម្រាប់ដំណើរការដង្ហើម និងរស្មីសំយោគ ក៏ដូចជាសម្រាប់ដំណើរការនៃអង់ស៊ីមជាច្រើន។
ធាតុដានមាននៅក្នុងរាងកាយក្នុងការប្រមូលផ្តុំតិចជាង 0.01% ហើយកំហាប់សរុបរបស់ពួកគេនៅក្នុងកោសិកាមិនឈានដល់ 0.1% ទេ។ ធាតុដានរួមមានស័ង្កសីទង់ដែងម៉ង់ហ្គាណែស cobalt អ៊ីយ៉ូត fluorine ជាដើម។ ស័ង្កសីគឺជាផ្នែកមួយនៃម៉ូលេគុលអរម៉ូនលំពែង អាំងស៊ុយលីន ទង់ដែងត្រូវបានទាមទារសម្រាប់ការធ្វើរស្មីសំយោគ និងការដកដង្ហើម។ Cobalt គឺជាសមាសធាតុនៃវីតាមីន B 12 ដែលអវត្តមានដែលនាំឱ្យមានភាពស្លេកស្លាំង។ អ៊ីយ៉ូតគឺចាំបាច់សម្រាប់ការសំយោគអរម៉ូនទីរ៉ូអ៊ីត ដែលធានានូវដំណើរការធម្មតានៃការរំលាយអាហារ ហើយហ្វ្លុយអូរីនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើតស្រទាប់ធ្មេញ។
ទាំងកង្វះ និងលើស ឬបញ្ហាមេតាបូលីសនៃ macro- និង microelements នាំឱ្យមានការវិវត្តនៃជំងឺផ្សេងៗ។ ជាពិសេសកង្វះជាតិកាល់ស្យូមនិងផូស្វ័របណ្តាលឱ្យ rickets កង្វះអាសូត - កង្វះប្រូតេអ៊ីនធ្ងន់ធ្ងរកង្វះជាតិដែក - ភាពស្លេកស្លាំងនិងកង្វះអ៊ីយ៉ូត - ការរំលោភលើការបង្កើតអរម៉ូនទីរ៉ូអ៊ីតនិងការថយចុះអត្រាមេតាប៉ូលីស។ ការកាត់បន្ថយការទទួលទានជាតិហ្វ្លុយអូរីជាមួយនឹងទឹក និងអាហារក្នុងកម្រិតធំបណ្តាលឱ្យមានការរំលោភលើការបន្តឡើងវិញនៃស្រទាប់ធ្មេញ ហើយជាលទ្ធផល បណ្តាលឱ្យមានហានិភ័យចំពោះសត្វ caries ។ សំណគឺពុលដល់សារពាង្គកាយស្ទើរតែទាំងអស់។ ការលើសរបស់វាបណ្តាលឱ្យខូចខាតជាអចិន្ត្រៃយ៍ដល់ខួរក្បាល និងប្រព័ន្ធសរសៃប្រសាទកណ្តាល ដែលត្រូវបានបង្ហាញដោយការបាត់បង់ការមើលឃើញ និងការស្តាប់ ការគេងមិនលក់ ខ្សោយតម្រងនោម ប្រកាច់ ហើយក៏អាចនាំឱ្យខ្វិន និងជំងឺដូចជាមហារីកផងដែរ។ ការពុលស្រួចស្រាវត្រូវបានអមដោយការយល់ឃើញភ្លាមៗ ហើយបញ្ចប់ដោយសន្លប់ និងស្លាប់។ 
អង្ករ។ ២.៤. ខ្លឹមសារនៃសារធាតុគីមីនៅក្នុងកោសិកា
កង្វះម៉ាក្រូ និងមីក្រូធាតុអាចត្រូវបានផ្តល់សំណងដោយការបង្កើនមាតិការបស់វានៅក្នុងអាហារ និងទឹកផឹក ក៏ដូចជាដោយការលេបថ្នាំ។ ដូច្នេះ អ៊ីយ៉ូតមាននៅក្នុងអាហារសមុទ្រ និងអំបិលអ៊ីយ៉ូត កាល់ស្យូមនៅក្នុងសំបកស៊ុត។ល។
២.៣.១. សារធាតុអសរីរាង្គនៃកោសិកា។
ធាតុគីមីនៃកោសិកាបង្កើតជាសមាសធាតុផ្សេងៗគ្នា - សរីរាង្គនិងសរីរាង្គ។ សារធាតុអសរីរាង្គនៃកោសិការួមមានទឹក អំបិលរ៉ែ អាស៊ីត។ល។ ហើយសារធាតុសរីរាង្គរួមមាន ប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត lipid ATP វីតាមីន។ល។ (រូបភាព 2.4)។
ទឹក (H 2 0) គឺជាសារធាតុអសរីរាង្គទូទៅបំផុតនៃកោសិកា ដែលមានលក្ខណៈសម្បត្តិរូបវិទ្យាតែមួយគត់។ វាគ្មានរសជាតិ គ្មានពណ៌ គ្មានក្លិន។ ដង់ស៊ីតេ និង viscosity នៃសារធាតុទាំងអស់ត្រូវបានប៉ាន់ស្មានដោយទឹក។ ដូចសារធាតុផ្សេងទៀតដែរ ទឹកអាចស្ថិតនៅក្នុងស្ថានភាពបីនៃការប្រមូលផ្តុំ៖ រឹង (ទឹកកក) រាវ និងឧស្ម័ន (ចំហាយទឹក)។ ចំណុចរលាយនៃទឹកគឺ 0°C ចំណុចរំពុះគឺ 100°C ទោះបីជាយ៉ាងណាក៏ដោយ ការរលាយនៃសារធាតុផ្សេងទៀតនៅក្នុងទឹកអាចផ្លាស់ប្តូរលក្ខណៈទាំងនេះបាន។ សមត្ថភាពកំដៅនៃទឹកក៏ខ្ពស់ផងដែរ - 4200 kJ / mol ។ K ដែលផ្តល់ឱ្យនាងនូវឱកាសដើម្បីចូលរួមក្នុងដំណើរការនៃ thermoregulation ។ នៅក្នុងម៉ូលេគុលទឹក អាតូមអ៊ីដ្រូសែនមានទីតាំងនៅមុំ 105 ° ខណៈពេលដែលគូអេឡិចត្រុងធម្មតាត្រូវបានទាញចេញដោយអាតូមអុកស៊ីហ្សែនដែលមានលក្ខណៈអេឡិចត្រុងច្រើនជាង។ នេះកំណត់លក្ខណៈសម្បត្តិ dipole នៃម៉ូលេគុលទឹក (ចុងម្ខាងរបស់ពួកគេត្រូវបានចោទប្រកាន់ជាវិជ្ជមាន ហើយមួយទៀតត្រូវបានចោទប្រកាន់អវិជ្ជមាន) និងលទ្ធភាពនៃការបង្កើតចំណងអ៊ីដ្រូសែនរវាងម៉ូលេគុលទឹក (រូបភាព 2.5) ។ ការស្អិតជាប់នៃម៉ូលេគុលទឹក បញ្ជាក់ពីបាតុភូតនៃភាពតានតឹងលើផ្ទៃ capillarity និងលក្ខណៈសម្បត្តិនៃទឹកជាសារធាតុរំលាយសកល។ ជាលទ្ធផលសារធាតុទាំងអស់ត្រូវបានបែងចែកទៅជា រលាយនៅក្នុងទឹក (hydrophilic) និង មិនរលាយនៅក្នុងវា (hydrophobic) ។ សូមអរគុណចំពោះលក្ខណៈសម្បត្តិពិសេសទាំងនេះ វាត្រូវបានគេកំណត់ទុកជាមុនថាទឹកបានក្លាយជាមូលដ្ឋាននៃជីវិតនៅលើផែនដី។
បរិមាណទឹកជាមធ្យមនៅក្នុងកោសិកានៃរាងកាយគឺមិនដូចគ្នាទេ ហើយអាចប្រែប្រួលទៅតាមអាយុ។ ដូច្នេះនៅក្នុងអំប្រ៊ីយ៉ុងរបស់មនុស្សដែលមានអាយុមួយខែកន្លះ បរិមាណទឹកនៅក្នុងកោសិកាឈានដល់ 97.5% ក្នុងអាយុប្រាំបីខែ - 83% ចំពោះទារកទើបនឹងកើតវាថយចុះដល់ 74% ហើយចំពោះមនុស្សពេញវ័យវាមានជាមធ្យម 66% ។ ទោះជាយ៉ាងណាក៏ដោយ កោសិការាងកាយមានភាពខុសគ្នានៅក្នុងមាតិកាទឹក។ ដូច្នេះឆ្អឹងមានទឹកប្រហែល 20% ថ្លើម - 70% និងខួរក្បាល - 86% ។ សរុបមក វាអាចនិយាយបានថា ការប្រមូលផ្តុំទឹកនៅក្នុងកោសិកាគឺសមាមាត្រដោយផ្ទាល់ទៅនឹងអត្រាមេតាប៉ូលីស។
អំបិលរ៉ែអាចស្ថិតក្នុងសភាពរលាយ ឬមិនរលាយ។ អំបិលរលាយបំបែកទៅជា ions - cations និង anions ។ cations សំខាន់បំផុតគឺប៉ូតាស្យូមនិងអ៊ីយ៉ុងសូដ្យូមដែលជួយសម្រួលដល់ការផ្ទេរសារធាតុនៅទូទាំងភ្នាសនិងចូលរួមក្នុងការកើតឡើងនិងដំណើរការនៃកម្លាំងសរសៃប្រសាទមួយ; ក៏ដូចជាអ៊ីយ៉ុងកាល់ស្យូម ដែលចូលរួមក្នុងដំណើរការនៃការកន្ត្រាក់នៃសរសៃសាច់ដុំ និងការកកឈាម។ ម៉ាញ៉េស្យូមដែលជាផ្នែកមួយនៃក្លរ៉ូហ្វីល; ជាតិដែក ដែលជាផ្នែកមួយនៃប្រូតេអ៊ីនមួយចំនួន រួមទាំងអេម៉ូក្លូប៊ីន។ anions សំខាន់បំផុតគឺ phosphate anion ដែលជាផ្នែកមួយនៃ ATP និងអាស៊ីត nucleic និងសំណល់អាស៊ីត carbonic ដែលបន្ទន់ការប្រែប្រួលនៃ pH របស់ឧបករណ៍ផ្ទុក។ អ៊ីយ៉ុងនៃអំបិលរ៉ែផ្តល់ទាំងការជ្រៀតចូលនៃទឹកខ្លួនវាទៅក្នុងកោសិកា និងការរក្សារបស់វានៅក្នុងវា។ ប្រសិនបើកំហាប់អំបិលក្នុងបរិយាកាសទាបជាងនៅក្នុងកោសិកា នោះទឹកជ្រាបចូលទៅក្នុងកោសិកា។ ម្យ៉ាងទៀត អ៊ីយ៉ុងកំណត់លក្ខណៈសតិបណ្ដោះអាសន្ននៃ cytoplasm ពោលគឺ សមត្ថភាពរបស់វាក្នុងការរក្សា pH អាល់កាឡាំងបន្តិចបន្តួចនៃ cytoplasm ទោះបីជាមានការបង្កើតផលិតផលអាស៊ីត និងអាល់កាឡាំងថេរនៅក្នុងកោសិកាក៏ដោយ។

អំបិលមិនរលាយ(CaCO 3, Ca 3 (P0 4) 2 ។ល។) គឺជាផ្នែកមួយនៃឆ្អឹង ធ្មេញ សំបក និងសំបករបស់សត្វ unicellular និង multicellular
លើសពីនេះទៀត សមាសធាតុអសរីរាង្គផ្សេងទៀត ដូចជាអាស៊ីត និងអុកស៊ីដ អាចត្រូវបានផលិតនៅក្នុងសារពាង្គកាយ។ ដូច្នេះកោសិកា parietal នៃក្រពះរបស់មនុស្សផលិតអាស៊ីត hydrochloric ដែលធ្វើសកម្មភាពអង់ស៊ីម pepsin រំលាយអាហារហើយស៊ីលីកុនអុកស៊ីដ impregnates ជញ្ជាំងកោសិកានៃ horsetails និងបង្កើតជាសំបក diatom ។ ក្នុងប៉ុន្មានឆ្នាំថ្មីៗនេះតួនាទីរបស់នីទ្រីកអុកស៊ីដ (II) ក្នុងការផ្តល់សញ្ញានៅក្នុងកោសិកានិងរាងកាយក៏ត្រូវបានស៊ើបអង្កេតផងដែរ។
សារធាតុសរីរាង្គនៅក្នុងកោសិកាពួកវាបង្កើតបាន 20-30% នៃម៉ាសកោសិកា។ ទាំងនេះរួមមាន biopolymers - ប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត ខ្លាញ់ ATP ជាដើម។ ប្រភេទផ្សេងគ្នានៃកោសិកាមានបរិមាណផ្សេងគ្នានៃសមាសធាតុសរីរាង្គ។ កាបូអ៊ីដ្រាតស្មុគស្មាញ គ្របដណ្តប់លើកោសិការុក្ខជាតិ ប្រូតេអ៊ីន និងខ្លាញ់លើសនៅក្នុងសត្វ។ ទោះជាយ៉ាងណាក៏ដោយ ក្រុមនីមួយៗនៃសារធាតុសរីរាង្គនៅក្នុងប្រភេទកោសិកាណាមួយអនុវត្តមុខងារដូចខាងក្រោមៈ ការផ្តល់ថាមពល ធ្វើជាសម្ភារៈសំណង់ ផ្ទុកព័ត៌មាន។ល។ កំប្រុក។ក្នុងចំណោមសារធាតុសរីរាង្គនៃកោសិកា ប្រូតេអ៊ីនកាន់កាប់កន្លែងដំបូងក្នុងន័យបរិមាណ និងតម្លៃ។ នៅក្នុងសត្វពួកគេមាន 50% នៃម៉ាស់ស្ងួតនៃកោសិកា។ នៅក្នុងខ្លួនមនុស្សមានម៉ូលេគុលប្រូតេអ៊ីនជាច្រើនប្រភេទដែលខុសគ្នាពីគ្នាទៅវិញទៅមក និងពីប្រូតេអ៊ីននៃសារពាង្គកាយដទៃទៀត។ ថ្វីបើមានភាពខុសគ្នាច្រើន និងភាពស្មុគស្មាញនៃរចនាសម្ព័ន្ធក៏ដោយ ក៏ប្រូតេអ៊ីនត្រូវបានបង្កើតឡើងពីអាស៊ីដអាមីណូចំនួន 20៖ អាស៊ីតអាមីណូមានលក្ខណៈសម្បត្តិ amphoteric ដូច្នេះ ពួកគេទាក់ទងគ្នាទៅវិញទៅមក៖ 
ចំណង Peptide៖

នៅពេលបញ្ចូលគ្នា ម៉ូលេគុលបង្កើតបាន: dipeptide, tripeptide ឬ polypeptide ។ វាគឺជាសមាសធាតុនៃអាស៊ីតអាមីណូ 20 ឬច្រើនជាងនេះ។ លំដាប់នៃការផ្លាស់ប្តូរអាស៊ីតអាមីណូនៅក្នុងម៉ូលេគុលគឺមានភាពចម្រុះបំផុត។ វាអនុញ្ញាតឱ្យមានអត្ថិភាព  វ៉ារ្យ៉ង់ដែលខុសគ្នានៅក្នុងតម្រូវការនិងលក្ខណៈសម្បត្តិនៃម៉ូលេគុលប្រូតេអ៊ីន។ លំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងម៉ូលេគុលត្រូវបានគេហៅថារចនាសម្ព័ន្ធ។ បឋម - លីនេអ៊ែរ។ អនុវិទ្យាល័យ - វង់។ កម្រិតទីបី - globules ។ Quaternary - សមាគមនៃ globules (អេម៉ូក្លូប៊ីន) ។ ការបាត់បង់រចនាសម្ព័ន្ធដោយម៉ូលេគុលត្រូវបានគេហៅថា denaturation ។ វាបណ្តាលមកពីការផ្លាស់ប្តូរសីតុណ្ហភាព pH វិទ្យុសកម្ម។ ជាមួយនឹងផលប៉ះពាល់បន្តិចបន្តួចម៉ូលេគុលអាចស្ដារលក្ខណៈសម្បត្តិរបស់វា។ វាត្រូវបានគេប្រើក្នុងថ្នាំ (ថ្នាំអង់ទីប៊ីយោទិច) ។ មុខងាររបស់ប្រូតេអ៊ីននៅក្នុងកោសិកាមានភាពចម្រុះ។ សំខាន់បំផុតគឺការសាងសង់។ ប្រូតេអ៊ីនត្រូវបានចូលរួមនៅក្នុងការបង្កើតភ្នាសកោសិកាទាំងអស់នៅក្នុងសរីរាង្គ។ មុខងារកាតាលីករមានសារៈសំខាន់ខ្លាំងណាស់ - អង់ស៊ីមទាំងអស់គឺជាប្រូតេអ៊ីន។ មុខងារម៉ូទ័រត្រូវបានផ្តល់ដោយប្រូតេអ៊ីន contractile ។ ការដឹកជញ្ជូន - មាននៅក្នុងការភ្ជាប់ធាតុគីមីនិងផ្ទេរពួកវាទៅជាលិកា។ មុខងារការពារត្រូវបានផ្តល់ដោយប្រូតេអ៊ីនពិសេស - អង្គបដិប្រាណដែលបង្កើតឡើងនៅក្នុង leukocytes ។ ប្រូតេអ៊ីនបម្រើជាប្រភពថាមពល - ជាមួយនឹងការបំបែកពេញលេញនៃប្រូតេអ៊ីន 1 ក្រាម 11.6 kJ ត្រូវបានបញ្ចេញ។ កាបូអ៊ីដ្រាត។ទាំងនេះគឺជាសមាសធាតុកាបូន អ៊ីដ្រូសែន និងអុកស៊ីសែន។ តំណាងដោយជាតិស្ករ។ កោសិកាមានរហូតដល់ 5% ។ អ្នកមានបំផុត - កោសិការុក្ខជាតិ - រហូតដល់ 90% នៃម៉ាស (ដំឡូងបារាំង) ។ ពួកគេត្រូវបានបែងចែកជាសាមញ្ញនិងស្មុគស្មាញ។ សាមញ្ញ - monosugar (គ្លុយកូស) C 6 H 12 O 6, ស្ករទំពាំងបាយជូ, fructose ។ Disahara - (sucrose) C] 2 H 22 O 11 beet និងស្ករអំពៅ។ Polysugar (សែលុយឡូស ម្សៅ) (C 6 H 10 O 5) n. កាបូអ៊ីដ្រាតអនុវត្តមុខងារជាចម្បងនៃអគារ និងថាមពល។ នៅពេលដែលកាបូអ៊ីដ្រាត 1 ក្រាមត្រូវបានកត់សុី 17.6 kJ ត្រូវបានបញ្ចេញ។ ម្សៅ និង glycogen បម្រើជាទុនបម្រុងថាមពលរបស់កោសិកា។ លីពីត។ទាំងនេះគឺជាសារធាតុខ្លាញ់ និងសារធាតុខ្លាញ់នៅក្នុងកោសិកា។ ពួកវាជា esters នៃ glycerol និងអាស៊ីតឆ្អែត និង unsaturated ទម្ងន់ម៉ូលេគុលខ្ពស់។ ពួកវាអាចរឹងនិងរាវ - ប្រេង។ រុក្ខជាតិមាននៅក្នុងគ្រាប់ពូជពី 5-15% នៃសារធាតុស្ងួត។ មុខងារសំខាន់គឺថាមពល - នៅពេលដែល 1 ក្រាមនៃជាតិខ្លាញ់ត្រូវបានបំបែក 38.9 kJ ត្រូវបានបញ្ចេញ។ ខ្លាញ់គឺជាឃ្លាំងផ្ទុកសារធាតុចិញ្ចឹម។ ខ្លាញ់អនុវត្តមុខងារអគារគឺជាអ៊ីសូឡង់កំដៅដ៏ល្អ។ អាស៊ីតនុយក្លេអ៊ីក។ទាំងនេះគឺជាសមាសធាតុសរីរាង្គដ៏ស្មុគស្មាញ។ ពួកវាមាន C, H 2, O 2, N 2, P. ដែលមាននៅក្នុង nuclei និង cytoplasm ។
វ៉ារ្យ៉ង់ដែលខុសគ្នានៅក្នុងតម្រូវការនិងលក្ខណៈសម្បត្តិនៃម៉ូលេគុលប្រូតេអ៊ីន។ លំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងម៉ូលេគុលត្រូវបានគេហៅថារចនាសម្ព័ន្ធ។ បឋម - លីនេអ៊ែរ។ អនុវិទ្យាល័យ - វង់។ កម្រិតទីបី - globules ។ Quaternary - សមាគមនៃ globules (អេម៉ូក្លូប៊ីន) ។ ការបាត់បង់រចនាសម្ព័ន្ធដោយម៉ូលេគុលត្រូវបានគេហៅថា denaturation ។ វាបណ្តាលមកពីការផ្លាស់ប្តូរសីតុណ្ហភាព pH វិទ្យុសកម្ម។ ជាមួយនឹងផលប៉ះពាល់បន្តិចបន្តួចម៉ូលេគុលអាចស្ដារលក្ខណៈសម្បត្តិរបស់វា។ វាត្រូវបានគេប្រើក្នុងថ្នាំ (ថ្នាំអង់ទីប៊ីយោទិច) ។ មុខងាររបស់ប្រូតេអ៊ីននៅក្នុងកោសិកាមានភាពចម្រុះ។ សំខាន់បំផុតគឺការសាងសង់។ ប្រូតេអ៊ីនត្រូវបានចូលរួមនៅក្នុងការបង្កើតភ្នាសកោសិកាទាំងអស់នៅក្នុងសរីរាង្គ។ មុខងារកាតាលីករមានសារៈសំខាន់ខ្លាំងណាស់ - អង់ស៊ីមទាំងអស់គឺជាប្រូតេអ៊ីន។ មុខងារម៉ូទ័រត្រូវបានផ្តល់ដោយប្រូតេអ៊ីន contractile ។ ការដឹកជញ្ជូន - មាននៅក្នុងការភ្ជាប់ធាតុគីមីនិងផ្ទេរពួកវាទៅជាលិកា។ មុខងារការពារត្រូវបានផ្តល់ដោយប្រូតេអ៊ីនពិសេស - អង្គបដិប្រាណដែលបង្កើតឡើងនៅក្នុង leukocytes ។ ប្រូតេអ៊ីនបម្រើជាប្រភពថាមពល - ជាមួយនឹងការបំបែកពេញលេញនៃប្រូតេអ៊ីន 1 ក្រាម 11.6 kJ ត្រូវបានបញ្ចេញ។ កាបូអ៊ីដ្រាត។ទាំងនេះគឺជាសមាសធាតុកាបូន អ៊ីដ្រូសែន និងអុកស៊ីសែន។ តំណាងដោយជាតិស្ករ។ កោសិកាមានរហូតដល់ 5% ។ អ្នកមានបំផុត - កោសិការុក្ខជាតិ - រហូតដល់ 90% នៃម៉ាស (ដំឡូងបារាំង) ។ ពួកគេត្រូវបានបែងចែកជាសាមញ្ញនិងស្មុគស្មាញ។ សាមញ្ញ - monosugar (គ្លុយកូស) C 6 H 12 O 6, ស្ករទំពាំងបាយជូ, fructose ។ Disahara - (sucrose) C] 2 H 22 O 11 beet និងស្ករអំពៅ។ Polysugar (សែលុយឡូស ម្សៅ) (C 6 H 10 O 5) n. កាបូអ៊ីដ្រាតអនុវត្តមុខងារជាចម្បងនៃអគារ និងថាមពល។ នៅពេលដែលកាបូអ៊ីដ្រាត 1 ក្រាមត្រូវបានកត់សុី 17.6 kJ ត្រូវបានបញ្ចេញ។ ម្សៅ និង glycogen បម្រើជាទុនបម្រុងថាមពលរបស់កោសិកា។ លីពីត។ទាំងនេះគឺជាសារធាតុខ្លាញ់ និងសារធាតុខ្លាញ់នៅក្នុងកោសិកា។ ពួកវាជា esters នៃ glycerol និងអាស៊ីតឆ្អែត និង unsaturated ទម្ងន់ម៉ូលេគុលខ្ពស់។ ពួកវាអាចរឹងនិងរាវ - ប្រេង។ រុក្ខជាតិមាននៅក្នុងគ្រាប់ពូជពី 5-15% នៃសារធាតុស្ងួត។ មុខងារសំខាន់គឺថាមពល - នៅពេលដែល 1 ក្រាមនៃជាតិខ្លាញ់ត្រូវបានបំបែក 38.9 kJ ត្រូវបានបញ្ចេញ។ ខ្លាញ់គឺជាឃ្លាំងផ្ទុកសារធាតុចិញ្ចឹម។ ខ្លាញ់អនុវត្តមុខងារអគារគឺជាអ៊ីសូឡង់កំដៅដ៏ល្អ។ អាស៊ីតនុយក្លេអ៊ីក។ទាំងនេះគឺជាសមាសធាតុសរីរាង្គដ៏ស្មុគស្មាញ។ ពួកវាមាន C, H 2, O 2, N 2, P. ដែលមាននៅក្នុង nuclei និង cytoplasm ។  ក) DNA គឺជាប៉ូលីណុក្លេអូទីតជីវសាស្រ្តដែលមានខ្សែសង្វាក់ពីរនៃនុយក្លេអូទីត។ នុយក្លេអូទីត - មានមូលដ្ឋានអាសូតចំនួន 4: 2 purines - Adenine និង Valine, 2 pyrimedins Cytosine និង Guanine ក៏ដូចជាស្ករ - deoxyribose និងសំណល់អាស៊ីតផូស្វ័រ។ នៅក្នុងខ្សែសង្វាក់នីមួយៗ នុយក្លេអូទីតត្រូវបានតភ្ជាប់ដោយចំណង covalent ។ ខ្សែសង្វាក់នៃ nucleotides បង្កើតជា helices ។ helix នៃ DNA ដែលផ្ទុកដោយប្រូតេអ៊ីនបង្កើតជារចនាសម្ព័ន្ធ - ក្រូម៉ូសូម។ ខ) RNA គឺជាវត្ថុធាតុ polymer ដែលជាម៉ូណូមឺរដែលជានុយក្លេអូទីតនៅជិត DNA មូលដ្ឋានអាសូត - A, G, C. ជំនួសឱ្យ thymine មាន Uration ។ កាបូអ៊ីដ្រាតនៃ RNA គឺ ribose មានសំណល់នៃអាស៊ីតផូស្វ័រ។
ក) DNA គឺជាប៉ូលីណុក្លេអូទីតជីវសាស្រ្តដែលមានខ្សែសង្វាក់ពីរនៃនុយក្លេអូទីត។ នុយក្លេអូទីត - មានមូលដ្ឋានអាសូតចំនួន 4: 2 purines - Adenine និង Valine, 2 pyrimedins Cytosine និង Guanine ក៏ដូចជាស្ករ - deoxyribose និងសំណល់អាស៊ីតផូស្វ័រ។ នៅក្នុងខ្សែសង្វាក់នីមួយៗ នុយក្លេអូទីតត្រូវបានតភ្ជាប់ដោយចំណង covalent ។ ខ្សែសង្វាក់នៃ nucleotides បង្កើតជា helices ។ helix នៃ DNA ដែលផ្ទុកដោយប្រូតេអ៊ីនបង្កើតជារចនាសម្ព័ន្ធ - ក្រូម៉ូសូម។ ខ) RNA គឺជាវត្ថុធាតុ polymer ដែលជាម៉ូណូមឺរដែលជានុយក្លេអូទីតនៅជិត DNA មូលដ្ឋានអាសូត - A, G, C. ជំនួសឱ្យ thymine មាន Uration ។ កាបូអ៊ីដ្រាតនៃ RNA គឺ ribose មានសំណល់នៃអាស៊ីតផូស្វ័រ។

RNAs ពីរខ្សែគឺជាអ្នកបញ្ជូនព័ត៌មានហ្សែន។ ខ្សែតែមួយ - ផ្ទុកព័ត៌មានអំពីលំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីន។ មាន RNA ខ្សែតែមួយជាច្រើន៖ - Ribosomal - នុយក្លេអូទីត 3-5 ពាន់; - ព័ត៌មាន - 300-30000 nucleotides; - ការដឹកជញ្ជូន - 76-85 nucleotides ។ ការសំយោគប្រូតេអ៊ីនត្រូវបានអនុវត្តនៅលើ ribosomes ដោយមានការចូលរួមពី RNA គ្រប់ប្រភេទ។
សំណួរសាកល្បង
1. កោសិកា - សារពាង្គកាយមួយឬផ្នែករបស់វា? 2. សមាសភាពបឋមនៃកោសិកា។ 3. ទឹក និងសារធាតុរ៉ែ។ 4. សារធាតុសរីរាង្គនៃកោសិកា។ 5. ប្រូតេអ៊ីន។ 6. កាបូអ៊ីដ្រាតខ្លាញ់។ 7. DNA ។ 8. RNA ។
ប្រធានបទ 2.2 រចនាសម្ព័ន្ធ និងមុខងាររបស់ក្រឡា
សំណួរសាកល្បង
1. តើកម្រិតនៃការរៀបចំកោសិកាមានន័យដូចម្តេច? 2. លក្ខណៈនៃ prokaryotes និង eukaryotes ។ 3. រចនាសម្ព័ន្ធនៃ prokaryotes ។ 4. សរីរវិទ្យានៃ prokaryotes ។ 5. រចនាសម្ព័ន្ធនៃ eukaryotes ។ 6. រចនាសម្ព័ន្ធនិងមុខងារនៃស្នូល។ 7. Karyotype និងលក្ខណៈពិសេសរបស់វា។ 8. រចនាសម្ព័ននិងមុខងារនៃស្នូល។ ប្រធានបទ 2.2.1 Golgi complex, lysosomes, mitochondria,
ribosomes មជ្ឈមណ្ឌលកោសិកា; សរីរាង្គនៃចលនា
ស៊ីតូប្លាស្មា- នេះគឺជាបរិយាកាសពាក់កណ្តាលរាវខាងក្នុងនៃកោសិកាដែលដំណើរការជីវគីមីទាំងអស់កើតឡើង។ វាមានរចនាសម្ព័ន្ធ - សរីរាង្គនិងទំនាក់ទំនងរវាងពួកគេ។ សរីរាង្គមានរចនាសម្ព័ន្ធ និងអាកប្បកិរិយាទៀងទាត់ក្នុងកំឡុងពេលផ្សេងៗគ្នានៃជីវិតកោសិកា និងអនុវត្តមុខងារជាក់លាក់។ មានសរីរាង្គលក្ខណៈនៃកោសិកាទាំងអស់ - mitochondria, មជ្ឈមណ្ឌលកោសិកា, Golgi apparatus, ribosomes, ER, lysosomes ។ សរីរាង្គនៃចលនា - flagella និង cilia គឺជាលក្ខណៈនៃសារពាង្គកាយឯកតា។ សារធាតុផ្សេងៗត្រូវបានដាក់ក្នុង cytoplasm - ការដាក់បញ្ចូល។ ទាំងនេះគឺជារចនាសម្ព័ន្ធអចិន្ត្រៃយ៍ដែលកើតឡើងនៅក្នុងដំណើរការនៃជីវិត។ ការរួមបញ្ចូលក្រាស់គឺជា granules ការរួមបញ្ចូលរាវគឺជា vacuoles ។ ទំហំរបស់ពួកគេត្រូវបានកំណត់ដោយសកម្មភាពសំខាន់នៃកោសិកា។ មូលដ្ឋាននៃការរៀបចំរចនាសម្ព័ន្ធនៃកោសិកាគឺជាគោលការណ៍ភ្នាសនៃរចនាសម្ព័ន្ធ។ នេះមានន័យថាកោសិកាត្រូវបានសាងសង់ជាចម្បងនៃភ្នាស។ ភ្នាសទាំងអស់មានរចនាសម្ព័ន្ធស្រដៀងគ្នា។ គំរូដែលទទួលយកត្រូវបានចាត់ទុកថាជារចនាសម្ព័ន្ធវត្ថុធាតុរាវ៖ ភ្នាសត្រូវបានបង្កើតឡើងដោយលីពីតពីរជួរដែលម៉ូលេគុលប្រូតេអ៊ីនត្រូវបានជ្រមុជនៅជម្រៅខុសៗគ្នា។ ភ្នាស cytoplasmic ខាងក្រៅវាមានវត្តមាននៅក្នុងកោសិកាទាំងអស់ ហើយបំបែក cytoplasm ពីបរិយាកាសខាងក្រៅ បង្កើតបានជាផ្ទៃក្រឡា។ ផ្ទៃក្រឡាគឺខុសគ្នា លក្ខណៈសរីរវិទ្យារបស់វាខុសគ្នា។ កោសិកាមានកម្លាំងនិងភាពបត់បែនខ្ពស់។ នៅក្នុងភ្នាស cytoplasmic មានរន្ធញើសដែលឆ្លងកាត់ម៉ូលេគុលនៃសារធាតុកើតឡើង។ ការបញ្ចូលសារធាតុចូលទៅក្នុងកោសិកាគឺជាដំណើរការដែលទាមទារថាមពល។ ភ្នាសកោសិកាមានទ្រព្យសម្បត្តិពាក់កណ្តាល permeability ។ យន្តការសម្រាប់ semipermeability គឺ osmosis ។ បន្ថែមពីលើ osmosis សារធាតុគីមីនិងសារធាតុរឹងអាចចូលទៅក្នុងកោសិកាដោយសារតែការ protrusions - ទាំងនេះគឺជា pinocetosis និង phagocytosis ។ ភ្នាស cytoplasmic ក៏ផ្តល់នូវទំនាក់ទំនងរវាងកោសិកានៅក្នុងជាលិកានៃសារពាង្គកាយពហុកោសិកាដោយសារតែការផ្នត់ និងការរីកដុះដាលជាច្រើន។កោសិកាជាប្រព័ន្ធជីវសាស្រ្ត
ទ្រឹស្តីកោសិកាទំនើប បទប្បញ្ញត្តិចម្បងរបស់វា តួនាទីក្នុងការបង្កើតរូបភាពវិទ្យាសាស្ត្រធម្មជាតិទំនើបនៃពិភពលោក។ ការអភិវឌ្ឍចំណេះដឹងអំពីកោសិកា។ រចនាសម្ព័ន្ធកោសិកានៃសារពាង្គកាយគឺជាមូលដ្ឋាននៃការរួបរួមនៃពិភពសរីរាង្គដែលជាភស្តុតាងនៃទំនាក់ទំនងនៃធម្មជាតិរស់នៅ
ទ្រឹស្តីកោសិកាទំនើប បទប្បញ្ញត្តិចម្បងរបស់វា តួនាទីក្នុងការបង្កើតរូបភាពវិទ្យាសាស្ត្រធម្មជាតិទំនើបនៃពិភពលោក
គោលគំនិតជាមូលដ្ឋានមួយក្នុងជីវវិទ្យាទំនើបគឺគំនិតដែលថាសារពាង្គកាយមានជីវិតទាំងអស់មានរចនាសម្ព័ន្ធកោសិកា។ វិទ្យាសាស្រ្តទាក់ទងនឹងការសិក្សាអំពីរចនាសម្ព័ន្ធនៃកោសិកា សកម្មភាពសំខាន់របស់វា និងអន្តរកម្មជាមួយបរិស្ថាន។ cytologyឥឡូវនេះគេហៅជាទូទៅថាជាជីវវិទ្យាកោសិកា។ Cytology ជំពាក់រូបរាងរបស់វាទៅនឹងការបង្កើតទ្រឹស្តីកោសិកា (1838-1839, M. Schleiden, T. Schwann, បំពេញបន្ថែមនៅឆ្នាំ 1855 ដោយ R. Virchow)។
ទ្រឹស្តីកោសិកាគឺជាគំនិតទូទៅនៃរចនាសម្ព័ន្ធ និងមុខងារនៃកោសិកាដែលជាឯកតារស់នៅ ការបន្តពូជ និងតួនាទីរបស់ពួកគេក្នុងការបង្កើតសារពាង្គកាយពហុកោសិកា។
បទប្បញ្ញត្តិសំខាន់ៗនៃទ្រឹស្តីកោសិកា៖
- កោសិកាគឺជាឯកតានៃរចនាសម្ព័ន្ធ សកម្មភាពជីវិត ការលូតលាស់ និងការអភិវឌ្ឍនៃសារពាង្គកាយមានជីវិត - មិនមានជីវិតនៅខាងក្រៅកោសិកាទេ។
- ក្រឡាគឺជាប្រព័ន្ធតែមួយដែលមានធាតុជាច្រើនដែលភ្ជាប់គ្នាទៅវិញទៅមកដោយធម្មជាតិ ដែលតំណាងឱ្យការបង្កើតអាំងតេក្រាលជាក់លាក់មួយ។
- កោសិកានៃសារពាង្គកាយទាំងអស់គឺស្រដៀងគ្នានៅក្នុងសមាសភាពគីមីរចនាសម្ព័ន្ធនិងមុខងាររបស់វា។
- កោសិកាថ្មីត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែកកោសិកាមេ ("កោសិកាពីកោសិកា")។
- កោសិកានៃសារពាង្គកាយពហុកោសិកាបង្កើតជាជាលិកា ហើយសរីរាង្គត្រូវបានបង្កើតឡើងដោយជាលិកា។ ជីវិតរបស់សារពាង្គកាយទាំងមូលត្រូវបានកំណត់ដោយអន្តរកម្មនៃកោសិកាធាតុផ្សំរបស់វា។
- កោសិកានៃសារពាង្គកាយពហុកោសិកាមានសំណុំហ្សែនពេញលេញ ប៉ុន្តែខុសគ្នាពីគ្នាទៅវិញទៅមកនៅក្នុងក្រុមផ្សេងៗគ្នានៃហ្សែនដំណើរការសម្រាប់ពួកវា ដែលបណ្តាលឱ្យមានភាពចម្រុះផ្នែក morphological និងមុខងារនៃកោសិកា - ភាពខុសគ្នា។
សូមអរគុណដល់ការបង្កើតទ្រឹស្តីកោសិកា វាច្បាស់ណាស់ថាកោសិកាគឺជាអង្គភាពតូចបំផុតនៃជីវិត ដែលជាប្រព័ន្ធរស់នៅបឋមដែលមានសញ្ញា និងលក្ខណៈសម្បត្តិទាំងអស់នៃភាវៈរស់។ ការបង្កើតទ្រឹស្ដីកោសិកាបានក្លាយជាតម្រូវការជាមុនដ៏សំខាន់បំផុតសម្រាប់ការអភិវឌ្ឍទស្សនៈលើតំណពូជ និងភាពប្រែប្រួល ចាប់តាំងពីការកំណត់អត្តសញ្ញាណនៃធម្មជាតិ និងលំនាំដើមរបស់ពួកវាបានបង្ហាញពីភាពជាសកលនៃរចនាសម្ព័ន្ធនៃសារពាង្គកាយរស់នៅដោយជៀសមិនរួច។ ការបង្ហាញពីការរួបរួមនៃសមាសធាតុគីមី និងផែនការរចនាសម្ព័ន្ធនៃកោសិកាបានបម្រើជាកម្លាំងរុញច្រានសម្រាប់ការអភិវឌ្ឍគំនិតអំពីប្រភពដើមនៃសារពាង្គកាយមានជីវិត និងការវិវត្តន៍របស់វា។ លើសពីនេះ ប្រភពដើមនៃសារពាង្គកាយពហុកោសិកាពីកោសិកាតែមួយកំឡុងពេលបង្កើតអំប្រ៊ីយ៉ុងបានក្លាយជា dogma នៃអំប្រ៊ីយ៉ុងទំនើប។
ការអភិវឌ្ឍចំណេះដឹងអំពីកោសិកា
រហូតមកដល់សតវត្សទី 17 បុរសមិនបានដឹងអ្វីទាល់តែសោះអំពីរចនាសម្ព័ន្ធតូចៗនៃវត្ថុជុំវិញគាត់ ហើយយល់ឃើញពិភពលោកដោយភ្នែកទទេ។ ឧបករណ៍សម្រាប់សិក្សាមីក្រូពិភពលោក មីក្រូទស្សន៍ត្រូវបានបង្កើតឡើងប្រហែលនៅក្នុងឆ្នាំ 1590 ដោយមេកានិកជនជាតិហូឡង់ G. និង Z. Jansen ប៉ុន្តែភាពមិនល្អឥតខ្ចោះរបស់វាបានធ្វើឱ្យវាមិនអាចពិនិត្យមើលវត្ថុតូចៗបានគ្រប់គ្រាន់។ មានតែការបង្កើតនៅលើមូលដ្ឋានរបស់វានៃអ្វីដែលគេហៅថាមីក្រូទស្សន៍បរិវេណដោយ K. Drebbel (1572-1634) ដែលរួមចំណែកដល់ការរីកចម្រើននៅក្នុងតំបន់នេះ។
នៅឆ្នាំ 1665 រូបវិទូជនជាតិអង់គ្លេស R. Hooke (1635-1703) បានកែលម្អការរចនានៃមីក្រូទស្សន៍ និងបច្ចេកវិទ្យានៃការកិនកញ្ចក់ ហើយដោយចង់ធ្វើឱ្យប្រាកដថាគុណភាពរូបភាពមានភាពប្រសើរឡើង គាត់បានពិនិត្យមើលផ្នែកនៃឆ្នុក ធ្យូង និងរុក្ខជាតិរស់នៅក្រោម វា។ នៅលើផ្នែកនានា គាត់បានរកឃើញរន្ធញើសតូចបំផុតដែលស្រដៀងនឹង Honeycomb ហើយហៅថាកោសិកា (ពីឡាតាំង។ កោសិកាកោសិកា, កោសិកា) ។ វាគួរឱ្យចាប់អារម្មណ៍ក្នុងការកត់សម្គាល់ថា R. Hooke បានចាត់ទុកភ្នាសកោសិកាជាធាតុផ្សំសំខាន់នៃកោសិកា។
នៅពាក់កណ្តាលទីពីរនៃសតវត្សទី 17 ស្នាដៃរបស់មីក្រូទស្សន៍លេចធ្លោបំផុត M. Malpighi (1628-1694) និង N. Gru (1641-1712) បានបង្ហាញខ្លួនដែលបានរកឃើញរចនាសម្ព័ន្ធកោសិកានៃរុក្ខជាតិជាច្រើនផងដែរ។
ដើម្បីប្រាកដថាអ្វីដែលអ្នក R. Hooke និងអ្នកវិទ្យាសាស្ត្រផ្សេងទៀតបានឃើញជាការពិត ពាណិជ្ជករជនជាតិហូឡង់ A. van Leeuwenhoek ដែលមិនមានការអប់រំពិសេសនោះ បានបង្កើតការរចនាមីក្រូទស្សន៍ដោយឯករាជ្យ ដែលខុសពីមូលដ្ឋានដែលមានស្រាប់ ហើយបានកែលម្អការផលិតកែវថត។ បច្ចេកវិទ្យា។ នេះបានអនុញ្ញាតឱ្យគាត់សម្រេចបាននូវការកើនឡើងពី 275-300 ដង និងដើម្បីពិចារណាព័ត៌មានលម្អិតនៃរចនាសម្ព័ន្ធដែលមានលក្ខណៈបច្ចេកទេសមិនអាចចូលដំណើរការបានសម្រាប់អ្នកវិទ្យាសាស្ត្រផ្សេងទៀត។ A. van Leeuwenhoek ជាអ្នកសង្កេតការណ៍ដែលមិនអាចប្រៀបផ្ទឹមបាន៖ គាត់បានគូសវាសដោយប្រុងប្រយ័ត្ន និងពិពណ៌នាអំពីអ្វីដែលគាត់បានឃើញនៅក្រោមមីក្រូទស្សន៍ ប៉ុន្តែមិនបានស្វែងរកការពន្យល់នោះទេ។ គាត់បានរកឃើញសារពាង្គកាយឯកត្តជន រួមទាំងបាក់តេរី បានរកឃើញស្នូល ក្លរ៉ូផ្លាស្ទិច ការឡើងក្រាស់នៃជញ្ជាំងកោសិកានៅក្នុងកោសិការុក្ខជាតិ ប៉ុន្តែការរកឃើញរបស់គាត់អាចត្រូវបានគេវាយតម្លៃច្រើននៅពេលក្រោយ។
ការរកឃើញធាតុផ្សំនៃរចនាសម្ព័ន្ធផ្ទៃក្នុងនៃសារពាង្គកាយនៅពាក់កណ្តាលទីមួយនៃសតវត្សទី 19 បានធ្វើតាមពីមួយទៅមួយ ។ G. Mol សម្គាល់នៅក្នុងកោសិការុក្ខជាតិ និងវត្ថុរាវដែលមានជាតិទឹក - កោសិកាបឹង រកឃើញរន្ធញើស។ អ្នករុក្ខសាស្ត្រជនជាតិអង់គ្លេស R. Brown (1773-1858) បានរកឃើញស្នូលនៅក្នុងកោសិកាផ្កាអ័រគីដេនៅឆ្នាំ 1831 បន្ទាប់មកវាត្រូវបានគេរកឃើញនៅក្នុងកោសិការុក្ខជាតិទាំងអស់។ អ្នកវិទ្យាសាស្ត្រជនជាតិឆេក J. Purkinje (1787-1869) បានណែនាំពាក្យ "protoplasm" (1840) ដើម្បីសំដៅទៅលើមាតិកា gelatinous ពាក់កណ្តាលរាវនៃកោសិកាដែលគ្មានស្នូល។ អ្នករុក្ខសាស្ត្រជនជាតិបែលហ្ស៊ិក M. Schleiden (1804-1881) ជឿនលឿនជាងសហសម័យរបស់គាត់ទាំងអស់ ដែលសិក្សាពីការអភិវឌ្ឍន៍ និងភាពខុសគ្នានៃរចនាសម្ព័ន្ធកោសិកាផ្សេងៗនៃរុក្ខជាតិខ្ពស់ជាង បានបង្ហាញថាសារពាង្គកាយរុក្ខជាតិទាំងអស់មានប្រភពចេញពីកោសិកាតែមួយ។ គាត់ក៏បានចាត់ទុកសាកសព nucleolus រាងមូលនៅក្នុងស្នូលនៃកោសិកាខ្នាតខ្ទឹមបារាំង (1842)។
នៅឆ្នាំ 1827 អ្នកជំនាញផ្នែកអំប្រ៊ីយ៉ុងជនជាតិរុស្សី K. Baer បានរកឃើញស៊ុតរបស់មនុស្ស និងថនិកសត្វដទៃទៀត ដោយហេតុនេះបដិសេធគំនិតដែលថារាងកាយបានអភិវឌ្ឍទាំងស្រុងពី gametes បុរស។ លើសពីនេះទៀតគាត់បានបង្ហាញពីការបង្កើតសារពាង្គកាយសត្វពហុកោសិកាពីកោសិកាតែមួយ - ស៊ុតបង្កកំណើតក៏ដូចជាភាពស្រដៀងគ្នានៃដំណាក់កាលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងនៃសត្វពហុកោសិកាដែលបង្ហាញពីការរួបរួមនៃប្រភពដើមរបស់វា។ ពត៌មានដែលប្រមូលបាននៅពាក់កណ្តាលសតវត្សទី 19 ទាមទារអោយមានការយល់ឃើញជាទូទៅ ដែលបានក្លាយជាទ្រឹស្តីកោសិកា។ ជីវវិទ្យាជំពាក់រូបមន្តរបស់វាចំពោះអ្នកសត្វវិទ្យាជនជាតិអាឡឺម៉ង់ T. Schwann (1810-1882) ដែលផ្អែកលើទិន្នន័យផ្ទាល់ខ្លួនរបស់គាត់ និងការសន្និដ្ឋានរបស់ M. Schleiden លើការអភិវឌ្ឍន៍នៃរុក្ខជាតិ បានផ្តល់យោបល់ថាប្រសិនបើស្នូលមានវត្តមាននៅក្នុងការបង្កើតណាមួយដែលអាចមើលឃើញនៅក្រោមមីក្រូទស្សន៍។ បន្ទាប់មកការបង្កើតនេះគឺជាកោសិកា។ ដោយផ្អែកលើលក្ខណៈវិនិច្ឆ័យនេះ T. Schwann បានបង្កើតបទប្បញ្ញត្តិសំខាន់ៗនៃទ្រឹស្តីកោសិកា។
គ្រូពេទ្យជនជាតិអាឡឺម៉ង់ និងជាអ្នកព្យាបាលរោគ R. Virchow (1821-1902) បានណែនាំសំណើសំខាន់មួយទៀតនៅក្នុងទ្រឹស្តីនេះ៖ កោសិកាកើតឡើងដោយការបែងចែកកោសិកាដើម ពោលគឺកោសិកាត្រូវបានបង្កើតឡើងតែពីកោសិកា ("កោសិកាពីកោសិកា")។
ចាប់តាំងពីការបង្កើតទ្រឹស្តីកោសិកា គោលលទ្ធិនៃកោសិកាជាឯកតានៃរចនាសម្ព័ន្ធ មុខងារ និងការអភិវឌ្ឍន៍របស់សារពាង្គកាយត្រូវបានអភិវឌ្ឍជាបន្តបន្ទាប់។ នៅចុងសតវត្សរ៍ទី 19 ដោយសារភាពជឿនលឿននៃបច្ចេកវិទ្យាមីក្រូទស្សន៍រចនាសម្ព័ន្ធនៃកោសិកាត្រូវបានបញ្ជាក់ឱ្យច្បាស់លាស់សរីរាង្គត្រូវបានពិពណ៌នា - ផ្នែកនៃកោសិកាដែលបំពេញមុខងារផ្សេងៗវិធីសាស្រ្តសម្រាប់ការបង្កើតកោសិកាថ្មី (mitosis, meiosis) ។ ត្រូវបានសិក្សា ហើយសារៈសំខាន់ដ៏សំខាន់បំផុតនៃរចនាសម្ព័ន្ធកោសិកាក្នុងការផ្ទេរលក្ខណៈសម្បត្តិតំណពូជបានច្បាស់លាស់។ ការប្រើប្រាស់វិធីសាស្រ្តស្រាវជ្រាវរូបវិទ្យា និងគីមីចុងក្រោយបំផុត បានធ្វើឱ្យវាអាចស្វែងយល់ពីដំណើរការនៃការផ្ទុក និងការបញ្ជូនព័ត៌មានតំណពូជ ក៏ដូចជាសិក្សាពីរចនាសម្ព័ន្ធដ៏ល្អនៃរចនាសម្ព័ន្ធកោសិកានីមួយៗ។ ទាំងអស់នេះបានរួមចំណែកដល់ការបំបែកវិទ្យាសាស្ត្រនៃកោសិកាទៅជាសាខាឯករាជ្យនៃចំណេះដឹង - cytology.
រចនាសម្ព័ន្ធកោសិកានៃសារពាង្គកាយ ភាពស្រដៀងគ្នានៃរចនាសម្ព័ន្ធកោសិកានៃសារពាង្គកាយទាំងអស់ - មូលដ្ឋាននៃការរួបរួមនៃពិភពសរីរាង្គ ភស្តុតាងនៃទំនាក់ទំនងនៃធម្មជាតិរស់នៅ។
រាល់ភាវៈរស់ដែលគេស្គាល់បច្ចុប្បន្ន (រុក្ខជាតិ សត្វ ផ្សិត និងបាក់តេរី) មានរចនាសម្ព័ន្ធកោសិកា។ សូម្បីតែមេរោគដែលមិនមានរចនាសម្ព័ន្ធកោសិកាក៏អាចបន្តពូជបានតែនៅក្នុងកោសិកាប៉ុណ្ណោះ។ កោសិកាគឺជាអង្គភាពរចនាសម្ព័ន្ធ និងមុខងារបឋមនៃការរស់នៅ ដែលមានវត្តមាននៅក្នុងការបង្ហាញទាំងអស់របស់វា ជាពិសេស ការបំប្លែងសារជាតិមេតាបូលីស និងការបំប្លែងថាមពល ការរំលាយអាហារ ការលូតលាស់ និងការអភិវឌ្ឍន៍ ការបន្តពូជ និងការឆាប់ខឹង។ ក្នុងពេលជាមួយគ្នានេះ វាស្ថិតនៅក្នុងកោសិកាដែលព័ត៌មានតំណពូជត្រូវបានរក្សាទុក ដំណើរការ និងដឹង។
ទោះបីជាមានភាពចម្រុះនៃកោសិកាក៏ដោយ ផែនការរចនាសម្ព័ន្ធសម្រាប់ពួកវាគឺដូចគ្នា៖ ពួកគេទាំងអស់មាន ឧបករណ៍តំណពូជជ្រមុជក្នុង cytoplasmនិងកោសិកាជុំវិញ ភ្នាសប្លាស្មា.
កោសិកាបានកើតឡើងជាលទ្ធផលនៃការវិវត្តន៍ដ៏យូរនៃពិភពសរីរាង្គ។ ការបង្រួបបង្រួមកោសិកាទៅក្នុងសារពាង្គកាយពហុកោសិកា មិនមែនជាការបូកសរុបដ៏សាមញ្ញនោះទេ ព្រោះកោសិកានីមួយៗ ខណៈពេលដែលរក្សាបាននូវលក្ខណៈទាំងអស់ដែលមាននៅក្នុងសារពាង្គកាយមានជីវិត ក្នុងពេលជាមួយគ្នានោះ ទទួលបាននូវលក្ខណៈសម្បត្តិថ្មី ដោយសារតែដំណើរការនៃមុខងារជាក់លាក់មួយ។ ម្យ៉ាងវិញទៀត សារពាង្គកាយពហុកោសិកាអាចត្រូវបានបែងចែកទៅជាផ្នែកធាតុផ្សំរបស់វា - កោសិកា ប៉ុន្តែម្យ៉ាងវិញទៀត ការដាក់ពួកវារួមគ្នាម្តងទៀត វាមិនអាចទៅរួចទេក្នុងការស្តារមុខងាររបស់សារពាង្គកាយអាំងតេក្រាល ចាប់តាំងពីលក្ខណៈសម្បត្តិថ្មីលេចឡើងតែក្នុងអន្តរកម្មនៃ ផ្នែកនៃប្រព័ន្ធ។ នេះបង្ហាញពីគំរូសំខាន់មួយដែលកំណត់លក្ខណៈនៃការរស់នៅ ការរួបរួមនៃការមិនដាច់ពីគ្នា និងអាំងតេក្រាល។ ទំហំតូច និងកោសិកាមួយចំនួនធំបង្កើតផ្ទៃធំក្នុងសារពាង្គកាយពហុកោសិកា ដែលចាំបាច់ដើម្បីធានាបាននូវការរំលាយអាហារលឿន។ លើសពីនេះទៀតនៅក្នុងព្រឹត្តិការណ៍នៃការស្លាប់នៃផ្នែកមួយនៃរាងកាយភាពសុចរិតរបស់វាអាចត្រូវបានស្ដារឡើងវិញដោយសារតែការបន្តពូជនៃកោសិកា។ នៅខាងក្រៅកោសិកាការផ្ទុកនិងការបញ្ជូនព័ត៌មានតំណពូជការផ្ទុកនិងការផ្ទេរថាមពលជាមួយនឹងការផ្លាស់ប្តូរជាបន្តបន្ទាប់របស់វាទៅជាការងារគឺមិនអាចទៅរួចទេ។ ជាចុងក្រោយ ការបែងចែកមុខងាររវាងកោសិកាក្នុងសារពាង្គកាយពហុកោសិកាបានផ្តល់ឱកាសច្រើនសម្រាប់សារពាង្គកាយដើម្បីសម្របខ្លួនទៅនឹងបរិយាកាសរបស់ពួកគេ និងជាតម្រូវការជាមុនសម្រាប់ភាពស្មុគស្មាញនៃអង្គការរបស់ពួកគេ។
ដូច្នេះការបង្កើតការរួបរួមនៃផែនការនៃរចនាសម្ព័ន្ធនៃកោសិកានៃសារពាង្គកាយមានជីវិតទាំងអស់បានបម្រើជាភស្តុតាងនៃការរួបរួមនៃប្រភពដើមនៃជីវិតទាំងអស់នៅលើផែនដី។
ភាពខុសគ្នានៃកោសិកា។ កោសិកា Prokaryotic និង eukaryotic ។ លក្ខណៈប្រៀបធៀបនៃកោសិកានៃរុក្ខជាតិ សត្វ បាក់តេរី ផ្សិត ភាពចម្រុះនៃកោសិកា
យោងតាមទ្រឹស្ដីកោសិកា កោសិកាមួយគឺជាអង្គភាពរចនាសម្ព័ន្ធ និងមុខងារតូចបំផុតនៃសារពាង្គកាយ ដែលមានលក្ខណៈសម្បត្តិទាំងអស់នៃភាវៈរស់។ យោងតាមចំនួនកោសិកា សារពាង្គកាយត្រូវបានបែងចែកទៅជា unicellular និង multicellular ។ កោសិកានៃសារពាង្គកាយឯកត្តជនមានជាសារពាង្គកាយឯករាជ្យ និងអនុវត្តមុខងារទាំងអស់នៃភាវៈរស់។ prokaryotes ទាំងអស់មានកោសិកាតែមួយ និង eukaryotes មួយចំនួន (ប្រភេទជាច្រើននៃសារាយ ផ្សិត និងប្រូហ្សូអា) ដែលធ្វើអោយមានការភ្ញាក់ផ្អើលជាមួយនឹងរូបរាង និងទំហំដ៏វិសេសវិសាល។ ទោះជាយ៉ាងណាក៏ដោយសារពាង្គកាយភាគច្រើននៅតែជាពហុកោសិកា។ កោសិការបស់ពួកគេមានឯកទេសដើម្បីបំពេញមុខងារជាក់លាក់ និងបង្កើតជាជាលិកា និងសរីរាង្គ ដែលមិនអាចត្រូវបានឆ្លុះបញ្ចាំងនៅក្នុងលក្ខណៈ morphological ។ ជាឧទាហរណ៍ រាងកាយរបស់មនុស្សត្រូវបានបង្កើតឡើងពីកោសិកាប្រហែល 10 14 ដែលតំណាងដោយប្រហែល 200 ប្រភេទដែលមានរូបរាង និងទំហំផ្សេងៗគ្នា។
រូបរាងនៃកោសិកាអាចជារាងមូល រាងស៊ីឡាំង គូប prismatic រាងឌីស fusiform stellate ជាដើម និង stellate - កោសិកានៃជាលិកាសរសៃប្រសាទ។ កោសិកាមួយចំនួនមិនមានរាងអចិន្ត្រៃយ៍ទាល់តែសោះ។ ទាំងនេះរួមបញ្ចូលជាដំបូងនៃការទាំងអស់ leukocytes ឈាម។
ទំហំកោសិកាក៏ប្រែប្រួលយ៉ាងខ្លាំងផងដែរ៖ កោសិកាភាគច្រើននៃសារពាង្គកាយពហុកោសិកាមានទំហំពី 10 ទៅ 100 មីក្រូន និងតូចបំផុត - 2-4 មីក្រូ។ ដែនកំណត់ទាបគឺដោយសារតែកោសិកាត្រូវតែមានសំណុំសារធាតុ និងរចនាសម្ព័ន្ធអប្បបរមា ដើម្បីធានាបាននូវសកម្មភាពសំខាន់ៗ ហើយទំហំកោសិកាធំពេកនឹងការពារការផ្លាស់ប្តូរសារធាតុ និងថាមពលជាមួយបរិស្ថាន ហើយនឹងរារាំងដំណើរការនៃការថែរក្សាផងដែរ។ homeostasis ។ ទោះយ៉ាងណាក៏ដោយ កោសិកាមួយចំនួនអាចមើលឃើញដោយភ្នែកទទេ។ ដំបូងបង្អស់ ទាំងនេះរួមបញ្ចូលកោសិកានៃផ្លែឈើរបស់ឪឡឹក និងដើមផ្លែប៉ោម ក៏ដូចជាស៊ុតរបស់ត្រី និងសត្វស្លាបផងដែរ។ ទោះបីជាវិមាត្រលីនេអ៊ែរមួយនៃក្រឡាលើសពីមធ្យមក៏ដោយ នៅសល់ទាំងអស់ត្រូវគ្នាទៅនឹងបទដ្ឋាន។ ឧទាហរណ៍ ការរីកដុះដាលនៃណឺរ៉ូនអាចលើសពីប្រវែង 1 ម៉ែត្រ ប៉ុន្តែអង្កត់ផ្ចិតរបស់វានឹងនៅតែត្រូវនឹងតម្លៃមធ្យម។ មិនមានទំនាក់ទំនងផ្ទាល់រវាងទំហំកោសិកា និងទំហំរាងកាយទេ។ ដូច្នេះកោសិកាសាច់ដុំរបស់ដំរី និងកណ្ដុរមានទំហំដូចគ្នា។
កោសិកា Prokaryotic និង eukaryotic
ដូចដែលបានរៀបរាប់ខាងលើ កោសិកាមានមុខងារស្រដៀងគ្នាជាច្រើន និងលក្ខណៈ morphological ។ ពួកវានីមួយៗមាន cytoplasm មួយដែលបានជ្រមុជនៅក្នុងវា។ ឧបករណ៍តំណពូជនិងបំបែកចេញពីបរិយាកាសខាងក្រៅ ភ្នាសប្លាស្មា, ឬ ប្លាស្មាដែលមិនរំខានដល់ដំណើរការរំលាយអាហារ និងថាមពល។ នៅខាងក្រៅភ្នាសកោសិកាក៏អាចមានជញ្ជាំងកោសិកាផងដែរ ដែលមានសារធាតុផ្សេងៗ ដែលបម្រើការពារកោសិកា និងជាប្រភេទគ្រោងឆ្អឹងខាងក្រៅ។
cytoplasm គឺជាមាតិកាទាំងមូលនៃកោសិកាដែលបំពេញចន្លោះរវាងភ្នាសប្លាស្មានិងរចនាសម្ព័ន្ធដែលមានព័ត៌មានហ្សែន។ វាមានសារធាតុសំខាន់ - hyaloplasm- និង organelles និងការរួមបញ្ចូលនៅក្នុងវា។ សរីរាង្គ- ទាំងនេះគឺជាសមាសធាតុអចិន្ត្រៃយ៍នៃកោសិកាដែលបំពេញមុខងារជាក់លាក់ ហើយការរួមបញ្ចូលគឺជាសមាសធាតុដែលលេចឡើង និងបាត់ក្នុងអំឡុងពេលជីវិតរបស់កោសិកា អនុវត្តមុខងារផ្ទុក ឬ excretory ជាចម្បង។ ការរួមបញ្ចូលជាញឹកញាប់ត្រូវបានបែងចែកទៅជារឹង និងរាវ។ ការរួមបញ្ចូលរឹងត្រូវបានតំណាងជាចម្បងដោយ granules និងអាចមានលក្ខណៈខុសគ្នា ខណៈពេលដែល vacuoles និងការធ្លាក់ចុះជាតិខ្លាញ់ត្រូវបានចាត់ទុកថាជាការរួមបញ្ចូលរាវ។

បច្ចុប្បន្ននេះមានកោសិកាសំខាន់ពីរប្រភេទគឺ prokaryotic និង eukaryotic ។
កោសិកា prokaryotic មិនមានស្នូលទេ ព័ត៌មានហ្សែនរបស់វាមិនត្រូវបានបំបែកចេញពី cytoplasm ដោយភ្នាសទេ។
តំបន់នៃ cytoplasm ដែលរក្សាទុកព័ត៌មានហ្សែននៅក្នុងកោសិកា prokaryotic ត្រូវបានគេហៅថា នុយក្លីអ៊ីត. នៅក្នុង cytoplasm នៃកោសិកា prokaryotic ប្រភេទមួយនៃ organelles ribosomes ត្រូវបានរកឃើញជាចម្បង ហើយសរីរាង្គដែលព័ទ្ធជុំវិញដោយភ្នាសគឺអវត្តមានទាំងស្រុង។ បាក់តេរីគឺជា prokaryotes ។
កោសិកា eukaryotic គឺជាកោសិកាដែលយ៉ាងហោចណាស់នៅដំណាក់កាលមួយនៃការអភិវឌ្ឍន៍ វាមាន ស្នូល- រចនាសម្ព័ន្ធពិសេសដែល DNA ស្ថិតនៅ។
cytoplasm នៃកោសិកា eukaryotic ត្រូវបានសម្គាល់ដោយភាពខុសគ្នាសំខាន់ៗនៃភ្នាស និងសរីរាង្គដែលមិនមែនជាភ្នាស។ សារពាង្គកាយ Eukaryotic រួមមានរុក្ខជាតិ សត្វ និងផ្សិត។ ទំហំនៃកោសិកា prokaryotic ជាក្បួនគឺជាលំដាប់នៃទំហំតូចជាងទំហំនៃកោសិកា eukaryotic ។ prokaryotes ភាគច្រើនគឺជាសារពាង្គកាយកោសិកាតែមួយ ចំណែក eukaryotes គឺពហុកោសិកា។
លក្ខណៈប្រៀបធៀបនៃរចនាសម្ព័ន្ធកោសិកានៃរុក្ខជាតិ សត្វ បាក់តេរី និងផ្សិត
បន្ថែមពីលើលក្ខណៈពិសេសនៃលក្ខណៈនៃ prokaryotes និង eukaryotes កោសិកានៃរុក្ខជាតិសត្វផ្សិតនិងបាក់តេរីមានលក្ខណៈពិសេសមួយចំនួនទៀត។ ដូច្នេះកោសិការុក្ខជាតិមានសរីរាង្គជាក់លាក់ - chloroplastsដែលកំណត់សមត្ថភាពរបស់ពួកគេក្នុងការធ្វើរស្មីសំយោគ ខណៈដែលនៅក្នុងសារពាង្គកាយផ្សេងទៀត សរីរាង្គទាំងនេះមិនត្រូវបានរកឃើញទេ។ ជាការពិតណាស់ នេះមិនមានន័យថាសារពាង្គកាយផ្សេងទៀតមិនមានលទ្ធភាពធ្វើរស្មីសំយោគទេ ចាប់តាំងពីឧទាហរណ៍នៅក្នុងបាក់តេរី វាកើតឡើងនៅលើការឈ្លានពាននៃប្លាស្មាម៉ា និងភ្នាសភ្នាសបុគ្គលនៅក្នុង cytoplasm ។
កោសិការុក្ខជាតិជាធម្មតាមានផ្ទុកនូវចន្លោះប្រហោងធំដែលពោរពេញទៅដោយកោសិកា។ នៅក្នុងកោសិការបស់សត្វ ផ្សិត និងបាក់តេរី ពួកគេក៏ត្រូវបានគេរកឃើញដែរ ប៉ុន្តែពួកគេមានប្រភពដើមខុសគ្នាទាំងស្រុង និងបំពេញមុខងារផ្សេងៗគ្នា។ សារធាតុបម្រុងសំខាន់ដែលរកឃើញក្នុងទម្រង់នៃការរួមបញ្ចូលរឹងគឺម្សៅនៅក្នុងរុក្ខជាតិ glycogen នៅក្នុងសត្វ និងផ្សិត និង glycogen ឬ volutin នៅក្នុងបាក់តេរី។
លក្ខណៈពិសេសប្លែកមួយទៀតនៃក្រុមនៃសារពាង្គកាយទាំងនេះគឺការរៀបចំឧបករណ៍លើផ្ទៃ៖ កោសិកានៃសារពាង្គកាយសត្វមិនមានជញ្ជាំងកោសិកាទេ ភ្នាសប្លាស្មារបស់ពួកវាត្រូវបានគ្របដណ្ដប់ដោយ glycocalyx ស្តើង ខណៈដែលនៅសល់ទាំងអស់មានវា។ នេះគឺអាចយល់បានទាំងស្រុង ចាប់តាំងពីវិធីដែលសត្វចិញ្ចឹមត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការចាប់យកភាគល្អិតអាហារនៅក្នុងដំណើរការនៃ phagocytosis ហើយវត្តមាននៃជញ្ជាំងកោសិកានឹងដកហូតពួកគេពីលទ្ធភាពនេះ។ លក្ខណៈគីមីនៃសារធាតុដែលបង្កើតជញ្ជាំងកោសិកាគឺមិនដូចគ្នាទេនៅក្នុងក្រុមផ្សេងៗនៃសារពាង្គកាយមានជីវិត៖ ប្រសិនបើនៅក្នុងរុក្ខជាតិវាជាកោសិកា បន្ទាប់មកនៅក្នុងផ្សិតវាគឺជា chitin ហើយនៅក្នុងបាក់តេរីវាគឺជា murein ។ លក្ខណៈប្រៀបធៀបនៃរចនាសម្ព័ន្ធនៃកោសិកានៃរុក្ខជាតិសត្វផ្សិតនិងបាក់តេរី
| សញ្ញា | បាក់តេរី | សត្វ | ផ្សិត | រុក្ខជាតិ |
| វិធីសាស្រ្តចិញ្ចឹម | heterotrophic ឬ autotrophic | Heterotrophic | Heterotrophic | អូតូត្រូហ្វីក |
| ការរៀបចំព័ត៌មានតំណពូជ | prokaryotes | eukaryotes | eukaryotes | eukaryotes |
| ការធ្វើមូលដ្ឋានីយកម្ម DNA | នុយក្លេអ៊ីត, ប្លាស្មា | ស្នូល, មីតូខនឌ្រី | ស្នូល, មីតូខនឌ្រី | នុយក្លេអ៊ែរ, មីតូខនឌ្រី, ផ្លាស្ទីត |
| ភ្នាសប្លាស្មា | មាន | មាន | មាន | មាន |
| ជញ្ជាំងកោសិកា | Mureinovaya | — | Chitinous | សែលុយឡូស |
| ស៊ីតូប្លាស្មា | មាន | មាន | មាន | មាន |
| សរីរាង្គ | រីបូសូម | Membrane និង non-membrane រួមទាំងមជ្ឈមណ្ឌលកោសិកា | ភ្នាស និងមិនមែនភ្នាស | Membrane និង non-membrane រួមទាំង plastids |
| សរីរាង្គនៃចលនា | Flagella និង villi | Flagella និង cilia | Flagella និង cilia | Flagella និង cilia |
| Vacuoles | កម្រណាស់។ | contractile, រំលាយអាហារ | ពេលខ្លះ | vacuole កណ្តាលជាមួយបឹងទន្លេសាប |
| ការរួមបញ្ចូល | គ្លីកូហ្សែន, វ៉ុលទីន | គ្លីកូហ្សែន | គ្លីកូហ្សែន | ម្សៅ |
ភាពខុសគ្នានៃរចនាសម្ព័ន្ធកោសិកានៃអ្នកតំណាងនៃនគរផ្សេងៗគ្នានៃសត្វព្រៃត្រូវបានបង្ហាញនៅក្នុងរូបភាព។

សមាសធាតុគីមីនៃកោសិកា។ ម៉ាក្រូ និងមីក្រូធាតុ។ ទំនាក់ទំនងនៃរចនាសម្ព័ន្ធ និងមុខងារនៃសារធាតុអសរីរាង្គ និងសរីរាង្គ (ប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត lipid ATP) ដែលបង្កើតជាកោសិកា។ តួនាទីរបស់សារធាតុគីមីនៅក្នុងកោសិកា និងរាងកាយមនុស្ស
សមាសធាតុគីមីនៃកោសិកា
នៅក្នុងសមាសភាពនៃសារពាង្គកាយមានជីវិត ភាគច្រើននៃធាតុគីមីនៃតារាងតាមកាលកំណត់នៃធាតុនៃ D. I. Mendeleev ដែលបានរកឃើញរហូតមកដល់បច្ចុប្បន្នត្រូវបានរកឃើញ។ ម្យ៉ាងវិញទៀត ពួកវាមិនមានធាតុតែមួយដែលមិនមាននៅក្នុងធម្មជាតិគ្មានជីវិតទេ ហើយម្យ៉ាងវិញទៀត ការប្រមូលផ្តុំរបស់ពួកគេនៅក្នុងរូបកាយនៃធម្មជាតិគ្មានជីវិត និងសារពាង្គកាយមានជីវិតមានភាពខុសគ្នាខ្លាំង។
ធាតុគីមីទាំងនេះបង្កើតជាសារធាតុអសរីរាង្គ និងសារធាតុសរីរាង្គ។ ទោះបីជាការពិតដែលថាសារធាតុអសរីរាង្គមានឥទ្ធិពលលើសារពាង្គកាយមានជីវិតក៏ដោយ វាគឺជាសារធាតុសរីរាង្គដែលកំណត់ពីភាពប្លែកនៃសមាសធាតុគីមីរបស់វា និងបាតុភូតនៃជីវិតជាទូទៅ ដោយសារពួកវាត្រូវបានសំយោគជាចម្បងដោយសារពាង្គកាយក្នុងដំណើរការនៃសកម្មភាពសំខាន់ និងដើរតួនាទីយ៉ាងសំខាន់ក្នុង ប្រតិកម្ម។

វិទ្យាសាស្រ្តទាក់ទងនឹងការសិក្សាអំពីសមាសធាតុគីមីនៃសារពាង្គកាយ និងប្រតិកម្មគីមីដែលកើតឡើងនៅក្នុងពួកវា។ ជីវគីមី។
វាគួរតែត្រូវបានកត់សម្គាល់ថាមាតិកានៃសារធាតុគីមីនៅក្នុងកោសិកានិងជាលិកាផ្សេងគ្នាអាចប្រែប្រួលយ៉ាងខ្លាំង។ ជាឧទាហរណ៍ ខណៈពេលដែលប្រូតេអ៊ីននាំមុខក្នុងចំណោមសមាសធាតុសរីរាង្គនៅក្នុងកោសិកាសត្វ កាបូអ៊ីដ្រាតលើសនៅក្នុងកោសិការុក្ខជាតិ។
| ធាតុគីមី | សំបកផែនដី | ទឹកសមុទ្រ | សារពាង្គកាយមានជីវិត |
| អូ | 49.2 | 85.8 | 65-75 |
| គ | 0.4 | 0.0035 | 15-18 |
| ហ | 1.0 | 10.67 | 8-10 |
| ន | 0.04 | 0.37 | 1.5-3.0 |
| ទំ | 0.1 | 0.003 | 0.20-1.0 |
| ស | 0.15 | 0.09 | 0.15-0.2 |
| ខេ | 2.35 | 0.04 | 0.15-0.4 |
| Ca | 3.25 | 0.05 | 0.04-2.0 |
| ក្ល | 0.2 | 0.06 | 0.05-0.1 |
| មីលីក្រាម | 2.35 | 0.14 | 0.02-0.03 |
| ណា | 2.4 | 1.14 | 0.02-0.03 |
| ហ្វេ | 4.2 | 0.00015 | 0.01-0.015 |
| Zn | < 0.01 | 0.00015 | 0.0003 |
| គ | < 0.01 | < 0.00001 | 0.0002 |
| ខ្ញុំ | < 0.01 | 0.000015 | 0.0001 |
| ច | 0.1 | 2.07 | 0.0001 |
ម៉ាក្រូ និងមីក្រូធាតុ
ធាតុគីមីប្រហែល 80 ត្រូវបានរកឃើញនៅក្នុងសារពាង្គកាយមានជីវិត ប៉ុន្តែមានតែ 27 ធាតុទាំងនេះប៉ុណ្ណោះដែលមានមុខងាររបស់វានៅក្នុងកោសិកា និងសារពាង្គកាយ។ ធាតុដែលនៅសេសសល់គឺមានវត្តមាននៅក្នុងបរិមាណដាន ហើយហាក់ដូចជាត្រូវបានបញ្ចូលតាមរយៈអាហារ ទឹក និងខ្យល់។ ខ្លឹមសារនៃធាតុគីមីនៅក្នុងរាងកាយប្រែប្រួលយ៉ាងខ្លាំង។ អាស្រ័យលើកំហាប់ពួកវាត្រូវបានបែងចែកទៅជា macronutrients និង microelements ។
ការផ្តោតអារម្មណ៍នីមួយៗ macronutrientsនៅក្នុងរាងកាយលើសពី 0.01% ហើយមាតិកាសរុបរបស់ពួកគេគឺ 99% ។ Macronutrients រួមមាន អុកស៊ីហ្សែន កាបូន អ៊ីដ្រូសែន អាសូត ផូស្វ័រ ស្ពាន់ធ័រ ប៉ូតាស្យូម កាល់ស្យូម សូដ្យូម ក្លរីន ម៉ាញេស្យូម និងជាតិដែក។ ធាតុទាំងបួនដំបូង (អុកស៊ីហ្សែន កាបូន អ៊ីដ្រូសែន និងអាសូត) ត្រូវបានគេហៅថាផងដែរ។ សរីរាង្គចាប់តាំងពីពួកវាគឺជាផ្នែកមួយនៃសមាសធាតុសរីរាង្គសំខាន់ៗ។ ផូស្វ័រ និងស្ពាន់ធ័រ ក៏ជាសមាសធាតុនៃសារធាតុសរីរាង្គមួយចំនួនផងដែរ ដូចជាប្រូតេអ៊ីន និងអាស៊ីតនុយក្លេអ៊ីក។ ផូស្វ័រមានសារៈសំខាន់សម្រាប់ការបង្កើតឆ្អឹង និងធ្មេញ។
បើគ្មានសារធាតុ macronutrients ដែលនៅសល់ មុខងារធម្មតារបស់រាងកាយគឺមិនអាចទៅរួចទេ។ ដូច្នេះប៉ូតាស្យូមសូដ្យូមនិងក្លរីនត្រូវបានចូលរួមនៅក្នុងដំណើរការនៃការរំភើបនៃកោសិកា។ ប៉ូតាស្យូមក៏ត្រូវការសម្រាប់អង់ស៊ីមជាច្រើនដើម្បីដំណើរការ និងរក្សាទឹកនៅក្នុងកោសិកា។ កាល់ស្យូមត្រូវបានរកឃើញនៅក្នុងជញ្ជាំងកោសិកានៃរុក្ខជាតិ ឆ្អឹង ធ្មេញ និងសំបក mollusk ហើយត្រូវបានទាមទារសម្រាប់ការកន្ត្រាក់សាច់ដុំ និងចលនាខាងក្នុង។ ម៉ាញ៉េស្យូមគឺជាសមាសធាតុនៃក្លរ៉ូហ្វីល - សារធាតុពណ៌ដែលធានាលំហូរនៃការធ្វើរស្មីសំយោគ។ វាក៏ចូលរួមក្នុងការសំយោគប្រូតេអ៊ីនផងដែរ។ ជាតិដែក បន្ថែមពីលើការជាផ្នែកមួយនៃអេម៉ូក្លូប៊ីន ដែលផ្ទុកអុកស៊ីហ្សែនក្នុងឈាម គឺចាំបាច់សម្រាប់ដំណើរការដង្ហើម និងរស្មីសំយោគ ក៏ដូចជាសម្រាប់ដំណើរការនៃអង់ស៊ីមជាច្រើន។
ធាតុដានមាននៅក្នុងរាងកាយក្នុងការប្រមូលផ្តុំតិចជាង 0.01% ហើយកំហាប់សរុបរបស់ពួកគេនៅក្នុងកោសិកាមិនឈានដល់ 0.1% ទេ។ ធាតុដានរួមមានស័ង្កសីទង់ដែងម៉ង់ហ្គាណែស cobalt អ៊ីយ៉ូត fluorine ជាដើម។ ស័ង្កសីគឺជាផ្នែកមួយនៃម៉ូលេគុលអរម៉ូនលំពែង អាំងស៊ុយលីន ទង់ដែងត្រូវបានទាមទារសម្រាប់ការធ្វើរស្មីសំយោគ និងការដកដង្ហើម។ Cobalt គឺជាសមាសធាតុនៃវីតាមីន B12 ដែលអវត្តមានដែលនាំឱ្យមានភាពស្លេកស្លាំង។ អ៊ីយ៉ូតគឺចាំបាច់សម្រាប់ការសំយោគអរម៉ូនទីរ៉ូអ៊ីត ដែលធានានូវដំណើរការធម្មតានៃការរំលាយអាហារ ហើយហ្វ្លុយអូរីនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើតស្រទាប់ធ្មេញ។
ទាំងកង្វះ និងលើស ឬបញ្ហាមេតាបូលីសនៃ macro- និង microelements នាំឱ្យមានការវិវត្តនៃជំងឺផ្សេងៗ។ ជាពិសេស កង្វះជាតិកាល់ស្យូម និងផូស្វ័រ បណ្តាលឱ្យ rickets កង្វះអាសូត បណ្តាលឱ្យកង្វះប្រូតេអ៊ីនធ្ងន់ធ្ងរ កង្វះជាតិដែក ធ្វើឱ្យមានភាពស្លេកស្លាំង និងកង្វះអ៊ីយ៉ូតបណ្តាលឱ្យមានការរំលោភលើការបង្កើតអរម៉ូនទីរ៉ូអ៊ីត និងការថយចុះអត្រាមេតាប៉ូលីស។ ការកាត់បន្ថយការទទួលទានជាតិហ្វ្លុយអូរីជាមួយនឹងទឹក និងអាហារក្នុងកម្រិតធំបណ្តាលឱ្យមានការរំលោភលើការបន្តឡើងវិញនៃស្រទាប់ធ្មេញ ហើយជាលទ្ធផល បណ្តាលឱ្យមានហានិភ័យចំពោះសត្វ caries ។ សំណគឺពុលដល់សារពាង្គកាយស្ទើរតែទាំងអស់។ ការលើសរបស់វាបណ្តាលឱ្យខូចខាតជាអចិន្ត្រៃយ៍ដល់ខួរក្បាល និងប្រព័ន្ធសរសៃប្រសាទកណ្តាល ដែលត្រូវបានបង្ហាញដោយការបាត់បង់ការមើលឃើញ និងការស្តាប់ ការគេងមិនលក់ ខ្សោយតម្រងនោម ប្រកាច់ ហើយក៏អាចនាំឱ្យខ្វិន និងជំងឺដូចជាមហារីកផងដែរ។ ការពុលស្រួចស្រាវត្រូវបានអមដោយការយល់ឃើញភ្លាមៗ ហើយបញ្ចប់ដោយសន្លប់ និងស្លាប់។
កង្វះម៉ាក្រូ និងមីក្រូធាតុអាចត្រូវបានផ្តល់សំណងដោយការបង្កើនមាតិការបស់វានៅក្នុងអាហារ និងទឹកផឹក ក៏ដូចជាដោយការលេបថ្នាំ។ ដូច្នេះ អ៊ីយ៉ូតមាននៅក្នុងអាហារសមុទ្រ និងអំបិលអ៊ីយ៉ូត កាល់ស្យូមនៅក្នុងសំបកស៊ុត។ល។
ទំនាក់ទំនងនៃរចនាសម្ព័ន្ធ និងមុខងារនៃសារធាតុអសរីរាង្គ និងសរីរាង្គ (ប្រូតេអ៊ីន អាស៊ីត nucleic កាបូអ៊ីដ្រាត lipid ATP) ដែលបង្កើតជាកោសិកា។ តួនាទីរបស់សារធាតុគីមីនៅក្នុងកោសិកា និងរាងកាយមនុស្ស
សារធាតុអសរីរាង្គ
ធាតុគីមីនៃកោសិកាបង្កើតជាសមាសធាតុផ្សេងៗគ្នា - សរីរាង្គនិងសរីរាង្គ។ សារធាតុអសរីរាង្គនៃកោសិការួមមាន ទឹក អំបិលរ៉ែ អាស៊ីត។ល។ ហើយសារធាតុសរីរាង្គរួមមាន ប្រូតេអ៊ីន អាស៊ីតនុយក្លេអ៊ីក កាបូអ៊ីដ្រាត លីពីត ATP វីតាមីន។ល។
ទឹក។(H 2 O) - សារធាតុអសរីរាង្គទូទៅបំផុតនៃកោសិកាដែលមានលក្ខណៈសម្បត្តិរូបវិទ្យាតែមួយគត់។ វាគ្មានរសជាតិ គ្មានពណ៌ គ្មានក្លិន។ ដង់ស៊ីតេ និង viscosity នៃសារធាតុទាំងអស់ត្រូវបានប៉ាន់ស្មានដោយទឹក។ ដូចសារធាតុផ្សេងទៀតដែរ ទឹកអាចស្ថិតនៅក្នុងស្ថានភាពបីនៃការប្រមូលផ្តុំ៖ រឹង (ទឹកកក) រាវ និងឧស្ម័ន (ចំហាយទឹក)។ ចំណុចរលាយនៃទឹកគឺ $0°C ចំណុចរំពុះគឺ $100°C ទោះយ៉ាងណាក៏ដោយ ការរលាយនៃសារធាតុផ្សេងទៀតនៅក្នុងទឹកអាចផ្លាស់ប្តូរលក្ខណៈទាំងនេះបាន។ សមត្ថភាពកំដៅនៃទឹកក៏ខ្ពស់ផងដែរ - 4200 kJ / mol K ដែលធ្វើឱ្យវាអាចធ្វើទៅបានសម្រាប់វាចូលរួមក្នុងដំណើរការនៃ thermoregulation ។ នៅក្នុងម៉ូលេគុលទឹក អាតូមអ៊ីដ្រូសែនមានទីតាំងនៅមុំ $105°$ ខណៈពេលដែលគូអេឡិចត្រុងធម្មតាត្រូវបានទាញចេញដោយអាតូមអុកស៊ីហ្សែនដែលមានលក្ខណៈអេឡិចត្រុងច្រើនជាង។ នេះកំណត់លក្ខណៈសម្បត្តិ dipole នៃម៉ូលេគុលទឹក (ចុងម្ខាងរបស់វាត្រូវបានចោទប្រកាន់ជាវិជ្ជមាន ហើយមួយទៀតត្រូវបានចោទប្រកាន់អវិជ្ជមាន) និងលទ្ធភាពនៃការបង្កើតចំណងអ៊ីដ្រូសែនរវាងម៉ូលេគុលទឹក។ ការស្អិតជាប់នៃម៉ូលេគុលទឹក បញ្ជាក់ពីបាតុភូតនៃភាពតានតឹងលើផ្ទៃ capillarity និងលក្ខណៈសម្បត្តិនៃទឹកជាសារធាតុរំលាយសកល។ ជាលទ្ធផល សារធាតុទាំងអស់ត្រូវបានបែងចែកទៅជារលាយក្នុងទឹក (អ៊ីដ្រូហ្វីលីក) និងមិនរលាយក្នុងវា (អ៊ីដ្រូហ្វីលីក)។ សូមអរគុណចំពោះលក្ខណៈសម្បត្តិពិសេសទាំងនេះ វាត្រូវបានគេកំណត់ទុកជាមុនថាទឹកបានក្លាយជាមូលដ្ឋាននៃជីវិតនៅលើផែនដី។
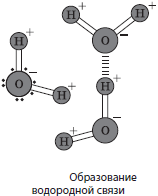
បរិមាណទឹកជាមធ្យមនៅក្នុងកោសិកានៃរាងកាយគឺមិនដូចគ្នាទេ ហើយអាចប្រែប្រួលទៅតាមអាយុ។ ដូច្នេះនៅក្នុងអំប្រ៊ីយ៉ុងរបស់មនុស្សដែលមានអាយុមួយខែកន្លះ បរិមាណទឹកនៅក្នុងកោសិកាឈានដល់ 97.5% ក្នុងអាយុប្រាំបីខែ - 83% ចំពោះទារកទើបនឹងកើតវាថយចុះដល់ 74% ហើយចំពោះមនុស្សពេញវ័យវាមានជាមធ្យម 66% ។ ទោះជាយ៉ាងណាក៏ដោយ កោសិការាងកាយមានភាពខុសគ្នានៅក្នុងមាតិកាទឹក។ ដូច្នេះឆ្អឹងមានទឹកប្រហែល 20% ថ្លើម - 70% និងខួរក្បាល - 86% ។ សរុបមក វាអាចនិយាយបានថា កំហាប់ទឹកនៅក្នុងកោសិកាគឺសមាមាត្រដោយផ្ទាល់ទៅនឹងអត្រាមេតាបូលីស.
អំបិលរ៉ែអាចស្ថិតក្នុងរដ្ឋរំលាយ ឬមិនទាន់រំលាយ។ អំបិលរលាយបំបែកទៅជា ions - cations និង anions ។ cations សំខាន់បំផុតគឺប៉ូតាស្យូមនិងអ៊ីយ៉ុងសូដ្យូមដែលជួយសម្រួលដល់ការផ្ទេរសារធាតុនៅទូទាំងភ្នាសនិងចូលរួមក្នុងការកើតឡើងនិងដំណើរការនៃកម្លាំងសរសៃប្រសាទមួយ; ក៏ដូចជាអ៊ីយ៉ុងកាល់ស្យូម ដែលចូលរួមក្នុងដំណើរការនៃការកន្ត្រាក់នៃសរសៃសាច់ដុំ និងការកកឈាម។ ម៉ាញ៉េស្យូមដែលជាផ្នែកមួយនៃក្លរ៉ូហ្វីល; ជាតិដែក ដែលជាផ្នែកមួយនៃប្រូតេអ៊ីនមួយចំនួន រួមទាំងអេម៉ូក្លូប៊ីន។ anions សំខាន់បំផុតគឺ phosphate anion ដែលជាផ្នែកមួយនៃ ATP និងអាស៊ីត nucleic និងសំណល់អាស៊ីត carbonic ដែលបន្ទន់ការប្រែប្រួលនៃ pH របស់ឧបករណ៍ផ្ទុក។ អ៊ីយ៉ុងនៃអំបិលរ៉ែផ្តល់ទាំងការជ្រៀតចូលនៃទឹកខ្លួនវាទៅក្នុងកោសិកា និងការរក្សារបស់វានៅក្នុងវា។ ប្រសិនបើកំហាប់អំបិលក្នុងបរិយាកាសទាបជាងនៅក្នុងកោសិកា នោះទឹកជ្រាបចូលទៅក្នុងកោសិកា។ ម្យ៉ាងទៀត អ៊ីយ៉ុងកំណត់លក្ខណៈសតិបណ្ដោះអាសន្ននៃ cytoplasm ពោលគឺ សមត្ថភាពរបស់វាក្នុងការរក្សា pH អាល់កាឡាំងបន្តិចបន្តួចនៃ cytoplasm ទោះបីជាមានការបង្កើតផលិតផលអាស៊ីត និងអាល់កាឡាំងថេរនៅក្នុងកោសិកាក៏ដោយ។
អំបិលមិនរលាយ(CaCO 3, Ca 3 (PO 4) 2 ។
លើសពីនេះទៀត សមាសធាតុអសរីរាង្គផ្សេងទៀត ដូចជាអាស៊ីត និងអុកស៊ីដ អាចត្រូវបានផលិតនៅក្នុងសារពាង្គកាយ។ ដូច្នេះកោសិកា parietal នៃក្រពះរបស់មនុស្សផលិតអាស៊ីត hydrochloric ដែលធ្វើសកម្មភាពអង់ស៊ីម pepsin រំលាយអាហារហើយស៊ីលីកុនអុកស៊ីដ impregnates ជញ្ជាំងកោសិកានៃ horsetails និងបង្កើតជាសំបក diatom ។ ក្នុងប៉ុន្មានឆ្នាំថ្មីៗនេះតួនាទីរបស់នីទ្រីកអុកស៊ីដ (II) ក្នុងការផ្តល់សញ្ញានៅក្នុងកោសិកានិងរាងកាយក៏ត្រូវបានស៊ើបអង្កេតផងដែរ។
បញ្ហាសរិរាង្គ
លក្ខណៈទូទៅនៃសារធាតុសរីរាង្គនៃកោសិកា
សារធាតុសរីរាង្គនៃកោសិកាអាចត្រូវបានតំណាងដោយទាំងម៉ូលេគុលសាមញ្ញ និងស្មុគស្មាញជាង។ ក្នុងករណីដែលម៉ូលេគុលស្មុគ្រស្មាញ (ម៉ាក្រូម៉ូលេគុល) ត្រូវបានបង្កើតឡើងដោយចំនួនដ៏សំខាន់នៃម៉ូលេគុលសាមញ្ញដែលកើតឡើងដដែលៗ វាត្រូវបានគេហៅថា វត្ថុធាតុ polymerនិងអង្គភាពរចនាសម្ព័ន្ធ - ម៉ូណូមឺរ. អាស្រ័យលើថាតើឯកតានៃប៉ូលីម៊ែរត្រូវបានធ្វើម្តងទៀតឬអត់នោះពួកគេត្រូវបានចាត់ថ្នាក់ជា ទៀងទាតឬ មិនទៀងទាត់. ប៉ូលីម័របង្កើតបានរហូតដល់ 90% នៃម៉ាស់សារធាតុស្ងួតនៃកោសិកា។ ពួកវាជាកម្មសិទ្ធិរបស់ថ្នាក់សំខាន់ៗចំនួនបីនៃសមាសធាតុសរីរាង្គ - កាបូអ៊ីដ្រាត (polysaccharides) ប្រូតេអ៊ីននិងអាស៊ីត nucleic ។ សារធាតុប៉ូលីម៊ែរធម្មតាគឺប៉ូលីសាខ័ររីត ខណៈដែលប្រូតេអ៊ីន និងអាស៊ីតនុយក្លេអ៊ីកមិនទៀងទាត់។ នៅក្នុងប្រូតេអ៊ីន និងអាស៊ីតនុយក្លេអ៊ីក លំដាប់នៃម៉ូណូមឺរមានសារៈសំខាន់ខ្លាំងណាស់ ព្រោះពួកវាបំពេញមុខងារព័ត៌មាន។

កាបូអ៊ីដ្រាត
កាបូអ៊ីដ្រាត- ទាំងនេះគឺជាសមាសធាតុសរីរាង្គ ដែលភាគច្រើនរួមបញ្ចូលធាតុគីមីចំនួនបី - កាបូន អ៊ីដ្រូសែន និងអុកស៊ីហ៊្សែន ទោះបីជាកាបូអ៊ីដ្រាតមួយចំនួនក៏មានផ្ទុកអាសូត ឬស្ពាន់ធ័រផងដែរ។ រូបមន្តទូទៅសម្រាប់កាបូអ៊ីដ្រាតគឺ C m (H 2 O) n ។ ពួកវាត្រូវបានបែងចែកទៅជាកាបូអ៊ីដ្រាតសាមញ្ញនិងស្មុគស្មាញ។
កាបូអ៊ីដ្រាតសាមញ្ញ (monosaccharides)មានម៉ូលេគុលជាតិស្ករតែមួយដែលមិនអាចបំបែកទៅជារបស់សាមញ្ញជាង។ ទាំងនេះគឺជាសារធាតុគ្រីស្តាល់ មានរសជាតិផ្អែម និងរលាយក្នុងទឹក។ Monosaccharides ចូលរួមយ៉ាងសកម្មក្នុងការរំលាយអាហារនៅក្នុងកោសិកា និងជាផ្នែកមួយនៃកាបូអ៊ីដ្រាតស្មុគស្មាញ - oligosaccharides និង polysaccharides ។
Monosaccharides ត្រូវបានចាត់ថ្នាក់ដោយចំនួនអាតូមកាបូន (C 3 -C 9) ឧទាហរណ៍។ pentoses(C 5) និង hexoses(ពី 6) ។ Pentoses រួមមាន ribose និង deoxyribose ។ រីបូសគឺជាផ្នែកមួយនៃ RNA និង ATP ។ Deoxyriboseគឺជាធាតុផ្សំនៃ DNA ។ Hexoses (C 6 H 12 O 6) គឺជាគ្លុយកូស, fructose, galactose ជាដើម។ គ្លុយកូស(ស្ករទំពាំងបាយជូ) ត្រូវបានរកឃើញនៅក្នុងសារពាង្គកាយទាំងអស់ រួមទាំងឈាមរបស់មនុស្សផងដែរ ព្រោះវាជាថាមពលបម្រុង។ វាគឺជាផ្នែកមួយនៃជាតិស្ករស្មុគស្មាញជាច្រើន: sucrose, lactose, maltose, ម្សៅ, សែលុយឡូសជាដើម។ Fructose(ស្ករផ្លែឈើ) ត្រូវបានគេរកឃើញនៅក្នុងការប្រមូលផ្តុំខ្ពស់បំផុតនៅក្នុងផ្លែឈើ ទឹកឃ្មុំ ដំណាំជា root beet ស្ករ។ វាមិនត្រឹមតែចូលរួមយ៉ាងសកម្មក្នុងដំណើរការមេតាបូលីសប៉ុណ្ណោះទេ ប៉ុន្តែវាក៏ជាផ្នែកនៃ sucrose និង polysaccharides មួយចំនួនដូចជាអាំងស៊ុយលីនផងដែរ។
monosaccharides ភាគច្រើនអាចផ្តល់ប្រតិកម្មកញ្ចក់ប្រាក់ និងកាត់បន្ថយទង់ដែងដោយបន្ថែមសារធាតុរាវរបស់ Fehling (ល្បាយនៃដំណោះស្រាយនៃទង់ដែង (II) sulfate និងប៉ូតាស្យូម-សូដ្យូម tartrate) ហើយដាំឱ្យពុះ។

TO oligosaccharidesរួមបញ្ចូលកាបូអ៊ីដ្រាតដែលបង្កើតឡើងដោយសំណល់ monosaccharide ជាច្រើន។ ជាទូទៅពួកវាក៏រលាយក្នុងទឹកខ្ពស់ និងមានរសជាតិផ្អែម។ អាស្រ័យលើចំនួនសំណល់ទាំងនេះ disaccharides (សំណល់ពីរ) trisaccharides (បី) ជាដើម។ Disaccharides រួមមាន sucrose, lactose, maltose ជាដើម។ sucrose(beet ឬស្ករអំពៅ) មានសំណល់នៃជាតិស្ករ និង fructose វាត្រូវបានរកឃើញនៅក្នុងសរីរាង្គផ្ទុករបស់រុក្ខជាតិមួយចំនួន។ ជាពិសេសជាច្រើននៃ sucrose នៅក្នុងឫសនៃ beet ស្ករនិងអំពៅដែលជាកន្លែងដែលពួកគេត្រូវបានទទួលបាននៅក្នុងវិធីឧស្សាហកម្មមួយ។ វាបម្រើជាគោលសម្រាប់ភាពផ្អែមនៃកាបូអ៊ីដ្រាត។ ឡាក់តូស, ឬ ស្ករទឹកដោះគោបង្កើតឡើងដោយសំណល់នៃជាតិគ្លុយកូស និងកាឡាក់តូស ដែលត្រូវបានរកឃើញនៅក្នុងទឹកដោះម្តាយ និងគោ។ ម៉ាល់តូស(ស្ករ malt) មានសំណល់គ្លុយកូសពីរ។ វាត្រូវបានបង្កើតឡើងកំឡុងពេលបំបែកសារធាតុ polysaccharides នៅក្នុងគ្រាប់ពូជរុក្ខជាតិ និងនៅក្នុងប្រព័ន្ធរំលាយអាហាររបស់មនុស្ស ហើយត្រូវបានគេប្រើក្នុងការផលិតស្រាបៀរ។

ប៉ូលីសាខារ៉ាតគឺជា biopolymers ដែល monomers គឺជា mono- ឬ disaccharide residue ។ សារធាតុ polysaccharides ភាគច្រើនមិនរលាយក្នុងទឹក ហើយរសជាតិមិនផ្អែម។ ទាំងនេះរួមមានម្សៅ, glycogen, cellulose និង chitin ។ ម្សៅ- នេះគឺជាសារធាតុម្សៅពណ៌ស ដែលមិនត្រូវបានសើមដោយទឹក ប៉ុន្តែបង្កើតបានជាការព្យួរនៅពេលញ៉ាំជាមួយទឹកក្តៅ - ម្សៅបិទភ្ជាប់។ ម្សៅពិតជាត្រូវបានបង្កើតឡើងពីប៉ូលីម៊ែរពីរ គឺអាមីឡូសដែលមានសាខាតិចជាង និងអាមីឡូផេកទីនដែលមានសាខាច្រើនជាង (រូបភាព 2.9)។ monomer ទាំង amylose និង amylopectin គឺគ្លុយកូស។ ម្សៅគឺជាសារធាតុផ្ទុកសំខាន់របស់រុក្ខជាតិ ដែលប្រមូលផ្តុំក្នុងបរិមាណដ៏ច្រើននៅក្នុងគ្រាប់ពូជ ផ្លែឈើ មើម មើម និងសរីរាង្គផ្ទុកផ្សេងទៀតរបស់រុក្ខជាតិ។ ប្រតិកម្មគុណភាពចំពោះម្សៅ គឺជាប្រតិកម្មជាមួយអ៊ីយ៉ូត ដែលម្សៅប្រែទៅជាពណ៌ខៀវ-ស្វាយ។

គ្លីកូហ្សែន(ម្សៅសត្វ) គឺជាសារធាតុ polysaccharide បម្រុងនៃសត្វ និងផ្សិត ដែលនៅក្នុងមនុស្សប្រមូលផ្តុំក្នុងបរិមាណដ៏ធំបំផុតនៅក្នុងសាច់ដុំ និងថ្លើម។ វាក៏មិនរលាយក្នុងទឹក និងមានរសជាតិមិនផ្អែម។ monomer នៃ glycogen គឺគ្លុយកូស។ បើប្រៀបធៀបទៅនឹងម៉ូលេគុលម្សៅ ម៉ូលេគុល glycogen គឺមានសាខាច្រើនជាង។
សែលុយឡូស, ឬ សែលុយឡូស, - polysaccharide យោងសំខាន់នៃរុក្ខជាតិ។ monomer នៃ cellulose គឺគ្លុយកូស។ ម៉ូលេគុលសែលុយឡូសដែលមិនមានសាខាបង្កើតជាបណ្តុំដែលជាផ្នែកមួយនៃជញ្ជាំងកោសិការបស់រុក្ខជាតិ។ សែលុយឡូសគឺជាមូលដ្ឋាននៃឈើវាត្រូវបានគេប្រើនៅក្នុងការសាងសង់ក្នុងការផលិតវាយនភ័ណ្ឌក្រដាសអាល់កុលនិងសារធាតុសរីរាង្គជាច្រើន។ សែលុយឡូសគឺអសកម្មគីមី ហើយមិនរលាយក្នុងអាស៊ីត ឬអាល់កាឡាំងទេ។ វាក៏មិនត្រូវបានបំបែកដោយអង់ស៊ីមនៃប្រព័ន្ធរំលាយអាហាររបស់មនុស្សដែរ ប៉ុន្តែបាក់តេរីនៅក្នុងពោះវៀនធំជួយរំលាយអាហារ។ លើសពីនេះទៀតជាតិសរសៃរំញោចការកន្ត្រាក់នៃជញ្ជាំងនៃការរលាក gastrointestinal នេះ, ជួយធ្វើឱ្យប្រសើរឡើងនូវការងាររបស់ខ្លួន។

ឈីទីនគឺជាសារធាតុប៉ូលីស្យូមឺរ ដែលជាម៉ូណូមឺរ ដែលជាសារធាតុ monosaccharide ដែលមានផ្ទុកអាសូត។ វាគឺជាផ្នែកមួយនៃជញ្ជាំងកោសិកានៃផ្សិត និងសែល arthropod ។ ក្នុងប្រព័ន្ធរំលាយអាហាររបស់មនុស្សក៏មិនមានអង់ស៊ីមសម្រាប់រំលាយសារធាតុ chitin ដែរ មានតែបាក់តេរីខ្លះប៉ុណ្ណោះដែលមានវា។
មុខងារនៃកាបូអ៊ីដ្រាត។កាបូអ៊ីដ្រាតអនុវត្តផ្លាស្ទិច (សំណង់) ថាមពលការផ្ទុកនិងមុខងារគាំទ្រនៅក្នុងកោសិកា។ ពួកវាបង្កើតជាជញ្ជាំងកោសិកានៃរុក្ខជាតិ និងផ្សិត។ តម្លៃថាមពលនៃការបំបែកកាបូអ៊ីដ្រាត 1 ក្រាមគឺ 17.2 kJ ។ គ្លុយកូស, fructose, sucrose, ម្សៅនិង glycogen គឺជាសារធាតុបម្រុង។ កាបូអ៊ីដ្រាតក៏អាចជាផ្នែកមួយនៃ lipids ស្មុគស្មាញនិងប្រូតេអ៊ីនបង្កើត glycolipids និង glycoproteins ជាពិសេសនៅក្នុងភ្នាសកោសិកា។ មិនមានសារៈសំខាន់តិចជាងនេះគឺតួនាទីនៃកាបូអ៊ីដ្រាតនៅក្នុងការទទួលស្គាល់អន្តរកោសិកានិងការយល់ឃើញនៃសញ្ញាបរិស្ថានចាប់តាំងពីពួកគេដើរតួជាអ្នកទទួលនៅក្នុងសមាសភាពនៃ glycoproteins ។
លីពីត
លីពីតគឺជាក្រុមគីមីនៃសារធាតុទម្ងន់ម៉ូលេគុលទាប ដែលមានលក្ខណៈសម្បត្តិ hydrophobic ។ សារធាតុទាំងនេះមិនរលាយក្នុងទឹក បង្កើតជាសារធាតុ emulsion នៅក្នុងវា ប៉ុន្តែងាយរលាយក្នុងសារធាតុរំលាយសរីរាង្គ។ Lipids មានជាតិខ្លាញ់នៅពេលប៉ះ ពួកវាភាគច្រើនបន្សល់ទុកនូវស្លាកស្នាមដែលមិនស្ងួតនៅលើក្រដាស។ រួមជាមួយនឹងប្រូតេអ៊ីន និងកាបូអ៊ីដ្រាត ពួកគេគឺជាសមាសធាតុសំខាន់មួយនៃកោសិកា។ ខ្លឹមសារនៃសារធាតុ lipids នៅក្នុងកោសិកាផ្សេងៗគ្នាគឺមិនដូចគ្នាទេ ជាពិសេសនៅក្នុងគ្រាប់ពូជ និងផ្លែឈើនៃរុក្ខជាតិមួយចំនួន នៅក្នុងថ្លើម បេះដូង និងឈាម។
អាស្រ័យលើរចនាសម្ព័ន្ធនៃម៉ូលេគុល lipid ត្រូវបានបែងចែកទៅជាសាមញ្ញ និងស្មុគស្មាញ។ TO សាមញ្ញ lipids រួមមាន lipids អព្យាក្រឹត (ខ្លាញ់) waxes និង steroids ។ ស្មុគស្មាញ lipid ក៏មានសមាសធាតុមួយទៀតដែលមិនមែនជាខ្លាញ់។ សំខាន់បំផុតក្នុងចំនោមពួកគេគឺ phospholipids glycolipids ជាដើម។
ខ្លាញ់គឺជា esters នៃ glycerol អាល់កុល trihydric និងអាស៊ីតខ្លាញ់ខ្ពស់ជាង។ អាស៊ីតខ្លាញ់ភាគច្រើនមានអាតូមកាបូន 14-22 ។ ក្នុងចំនោមពួកគេមានទាំង saturated និង unsaturated ពោលគឺមានចំណងទ្វេ។ ក្នុងចំណោមអាស៊ីតខ្លាញ់ឆ្អែត អាស៊ីត palmitic និង stearic គឺជារឿងធម្មតាបំផុត ហើយអាស៊ីតខ្លាញ់មិនឆ្អែតគឺ oleic ។ អាស៊ីតខ្លាញ់មិនឆ្អែតមួយចំនួនមិនត្រូវបានសំយោគនៅក្នុងខ្លួនមនុស្ស ឬត្រូវបានសំយោគក្នុងបរិមាណមិនគ្រប់គ្រាន់ទេ ដូច្នេះហើយមិនអាចខ្វះបាន។ សំណល់ glycerol បង្កើតជាក្បាល hydrophilic ខណៈដែលសំណល់អាស៊ីតខ្លាញ់បង្កើតជាកន្ទុយ hydrophobic ។

ខ្លាញ់អនុវត្តមុខងារផ្ទុកនៅក្នុងកោសិកា និងបម្រើជាប្រភពថាមពល។ ពួកវាសម្បូរទៅដោយជាលិកាខ្លាញ់ subcutaneous ដែលអនុវត្តមុខងារស្រូបទាញ និងអ៊ីសូឡង់កម្ដៅ ហើយនៅក្នុងសត្វក្នុងទឹក វាក៏បង្កើនភាពធន់ផងដែរ។ ខ្លាញ់រុក្ខជាតិភាគច្រើនមានអាស៊ីតខ្លាញ់មិនឆ្អែត ដែលជាលទ្ធផលដែលពួកវាជារាវ និងត្រូវបានគេហៅថា ប្រេង. ប្រេងមាននៅក្នុងគ្រាប់ពូជនៃរុក្ខជាតិជាច្រើនដូចជា ផ្កាឈូករ័ត្ន សណ្តែកសៀង រ៉េស ជាដើម។
ក្រមួនគឺជា esters និងល្បាយនៃអាស៊ីតខ្លាញ់ និងអាល់កុលខ្លាញ់។ នៅក្នុងរុក្ខជាតិពួកវាបង្កើតជាខ្សែភាពយន្តមួយនៅលើផ្ទៃស្លឹក ដែលការពារប្រឆាំងនឹងការហួត ការជ្រៀតចូលនៃភ្នាក់ងារបង្ករោគ។
TO ថ្នាំស្តេរ៉ូអ៊ីតរួមមាន lipid ដូចជា cholesterol ដែលជាសមាសធាតុសំខាន់នៃភ្នាសកោសិកា ក៏ដូចជាអរម៉ូនភេទ estradiol, testosterone, vitamin D ជាដើម។
ផូស្វ័រលីពីដបន្ថែមពីលើសំណល់នៃ glycerol និងអាស៊ីតខ្លាញ់ មានសំណល់នៃអាស៊ីត orthophosphoric ។ ពួកវាគឺជាផ្នែកមួយនៃភ្នាសកោសិកា និងផ្តល់នូវលក្ខណៈសម្បត្តិរារាំងរបស់ពួកគេ។
គ្លីកូលីពីដក៏ជាសមាសធាតុនៃភ្នាសដែរ ប៉ុន្តែខ្លឹមសាររបស់វាមានកម្រិតទាប។ ផ្នែកដែលមិនមានជាតិខ្លាញ់នៃ glycolipids គឺជាកាបូអ៊ីដ្រាត។
មុខងាររបស់ lipid ។ Lipids អនុវត្តផ្លាស្ទិច (ការកសាង) ថាមពល ការផ្ទុក ការការពារ ការបញ្ចេញ និងមុខងារនិយតកម្មនៅក្នុងកោសិកា លើសពីនេះទៀត ពួកគេគឺជាវីតាមីន។ វាគឺជាសមាសធាតុសំខាន់នៃភ្នាសកោសិកា។ នៅពេលពុះ 1 ក្រាមនៃ lipids ថាមពល 38.9 kJ ត្រូវបានបញ្ចេញ។ ពួកវាត្រូវបានតម្កល់នៅក្នុងទុនបំរុងនៅក្នុងសរីរាង្គផ្សេងៗនៃរុក្ខជាតិ និងសត្វ។ លើសពីនេះទៀតជាលិកាខ្លាញ់ក្រោមស្បែកការពារសរីរាង្គខាងក្នុងពីការថយចុះកម្តៅឬការឡើងកំដៅក៏ដូចជាការឆក់។ មុខងារនិយតកម្មនៃ lipid គឺដោយសារតែការពិតដែលថាពួកគេមួយចំនួនគឺជាអរម៉ូន។ រាងកាយខ្លាញ់នៃសត្វល្អិតបម្រើសម្រាប់ការ excretion ។
កំប្រុក
កំប្រុក- ទាំងនេះគឺជាសមាសធាតុម៉ូលេគុលខ្ពស់ ជីវប៉ូលីម័រ ម៉ូណូមឺរ ដែលជាអាស៊ីតអាមីណូដែលភ្ជាប់ដោយចំណង peptide ។
អាស៊ីតអាមីណូហៅថា សមាសធាតុសរីរាង្គ ដែលមានក្រុមអាមីណូ ក្រុម carboxyl និងរ៉ាឌីកាល់។ សរុបមក អាស៊ីដអាមីណូប្រហែល 200 ត្រូវបានរកឃើញនៅក្នុងធម្មជាតិ ដែលមានភាពខុសប្លែកគ្នានៅក្នុងរ៉ាឌីកាល់ និងការរៀបចំទៅវិញទៅមកនៃក្រុមមុខងារ ប៉ុន្តែមានតែ 20 ប៉ុណ្ណោះក្នុងចំណោមពួកវាអាចជាផ្នែកមួយនៃប្រូតេអ៊ីន។ អាស៊ីតអាមីណូទាំងនេះត្រូវបានគេហៅថា សារធាតុប្រូតេអ៊ីន។

ជាអកុសល មិនមែនអាស៊ីតអាមីណូប្រូតេអ៊ីនទាំងអស់អាចត្រូវបានសំយោគនៅក្នុងខ្លួនមនុស្សទេ ដូច្នេះពួកវាត្រូវបានបែងចែកទៅជាមិនសំខាន់ និងមិនអាចជំនួសបាន។ អាស៊ីតអាមីណូមិនសំខាន់ត្រូវបានបង្កើតឡើងនៅក្នុងខ្លួនមនុស្សក្នុងបរិមាណដែលត្រូវការ និង មិនអាចជំនួសបាន។- ទេ។ ពួកវាត្រូវតែមកពីអាហារ ប៉ុន្តែក៏អាចសំយោគដោយផ្នែកដោយអតិសុខុមប្រាណក្នុងពោះវៀនផងដែរ។ មានអាស៊ីដអាមីណូសំខាន់ៗចំនួន 8 យ៉ាង។ ទាំងនេះរួមមាន វ៉ាលីន អ៊ីសូលស៊ីន លីស៊ីន លីស៊ីន មេទីយ៉ូនីន ទ្រីអូនីន ទ្រីបតូហ្វាន និងហ្វីនីឡាឡានីន។ ទោះបីជាការពិតដែលថាអាស៊ីតអាមីណូប្រូតេអ៊ីនទាំងអស់ត្រូវបានសំយោគនៅក្នុងរុក្ខជាតិក៏ដោយក៏ប្រូតេអ៊ីនបន្លែមិនពេញលេញទេព្រោះវាមិនមានសំណុំអាស៊ីតអាមីណូពេញលេញលើសពីនេះទៅទៀតវត្តមាននៃប្រូតេអ៊ីននៅក្នុងផ្នែកលូតលាស់នៃរុក្ខជាតិកម្រមានលើសពី 1-2% នៃ ម៉ាស។ ដូច្នេះចាំបាច់ត្រូវញ៉ាំប្រូតេអ៊ីនមិនត្រឹមតែបន្លែប៉ុណ្ណោះទេប៉ុន្តែក៏មានប្រភពដើមពីសត្វផងដែរ។
លំដាប់នៃអាស៊ីតអាមីណូពីរដែលភ្ជាប់ដោយចំណង peptide ត្រូវបានគេហៅថា dipeptideក្នុងចំណោមបី tripeptideល ក្នុងចំណោម peptides មានសមាសធាតុសំខាន់ៗដូចជា អរម៉ូន (oxytocin, vasopressin) ថ្នាំអង់ទីប៊ីយោទិច។ល។ ខ្សែសង្វាក់នៃអាស៊ីតអាមីណូជាងម្ភៃត្រូវបានគេហៅថា polypeptideនិង polypeptides ដែលមានសំណល់អាស៊ីតអាមីណូច្រើនជាង 60 គឺជាប្រូតេអ៊ីន។

កម្រិតនៃអង្គការរចនាសម្ព័ន្ធប្រូតេអ៊ីន។ប្រូតេអ៊ីនអាចមានរចនាសម្ព័ន្ធបឋម អនុវិទ្យាល័យ ឧត្តមសិក្សា និងបួនជ្រុង។
រចនាសម្ព័ន្ធបឋមនៃប្រូតេអ៊ីន- នេះ។ លំដាប់អាស៊ីតអាមីណូលីនេអ៊ែរភ្ជាប់ដោយចំណង peptide ។ រចនាសម្ព័នបឋមកំណត់ពីភាពជាក់លាក់នៃប្រូតេអ៊ីន និងលក្ខណៈពិសេសរបស់វា ពីព្រោះទោះបីជាយើងសន្មត់ថាប្រូតេអ៊ីនជាមធ្យមមានសំណល់អាស៊ីតអាមីណូ 500 ក៏ដោយ នោះចំនួននៃបន្សំដែលអាចធ្វើបានគឺ 20,500។ ដូច្នេះហើយ ការផ្លាស់ប្តូរទីតាំងនៃអាមីណូយ៉ាងហោចណាស់មួយ អាស៊ីតនៅក្នុងរចនាសម្ព័ន្ធបឋមមានការផ្លាស់ប្តូររចនាសម្ព័ន្ធបន្ទាប់បន្សំ និងខ្ពស់ជាង ក៏ដូចជាលក្ខណៈសម្បត្តិនៃប្រូតេអ៊ីនទាំងមូល។

លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីនកំណត់ការវេចខ្ចប់របស់វា - ការលេចឡើងនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំនិងទីបី។
រចនាសម្ព័ន្ធបន្ទាប់បន្សំគឺជាការរៀបចំលំហនៃម៉ូលេគុលប្រូតេអ៊ីនក្នុងទម្រង់ វង់ឬ បត់រក្សាដោយចំណងអ៊ីដ្រូសែនរវាងអាតូមអុកស៊ីសែន និងអ៊ីដ្រូសែននៃក្រុម peptide នៃវេនផ្សេងគ្នានៃ helix ឬផ្នត់។ ប្រូតេអ៊ីនជាច្រើនមានតំបន់វែងច្រើនឬតិចជាមួយនឹងរចនាសម្ព័ន្ធបន្ទាប់បន្សំ។ ទាំងនេះគឺជាឧទាហរណ៍ keratins នៃសក់និងក្រចក, fibroin សូត្រ។

រចនាសម្ព័ន្ធទីបីកំប្រុក ( សកល) ក៏ជាទម្រង់នៃការបត់ជាលំហនៃខ្សែសង្វាក់ polypeptide ដែលផ្ទុកដោយ hydrophobic អ៊ីដ្រូសែន disulfide (S-S) និងចំណងផ្សេងទៀត។ វាគឺជាលក្ខណៈនៃប្រូតេអ៊ីនរាងកាយភាគច្រើនដូចជា myoglobin សាច់ដុំ។

រចនាសម្ព័ន្ធបួនជ្រុង- ស្មុគ្រស្មាញបំផុត បង្កើតឡើងដោយខ្សែសង្វាក់ polypeptide ជាច្រើនដែលតភ្ជាប់ជាចម្បងដោយចំណងដូចគ្នាដូចជានៅក្នុង tertiary (hydrophobic, ionic និង hydrogen) ក៏ដូចជាអន្តរកម្មខ្សោយផ្សេងទៀត។ រចនាសម្ព័ន្ធ quaternary គឺជាលក្ខណៈនៃប្រូតេអ៊ីនមួយចំនួនដូចជា អេម៉ូក្លូប៊ីន ក្លរ៉ូហ្វីល ជាដើម។

រូបរាងរបស់ម៉ូលេគុលគឺ fibrillarនិង រាងមូលប្រូតេអ៊ីន។ ទីមួយនៃពួកវាត្រូវបានពន្លូត ដូចជាឧទាហរណ៍ ជាលិកាភ្ជាប់ collagen ឬសក់ និងក្រចក keratins ។ ប្រូតេអ៊ីន Globular មានទម្រង់ជាបាល់ (globules) ដូចជាសាច់ដុំ myoglobin ។
ប្រូតេអ៊ីនសាមញ្ញនិងស្មុគស្មាញ។ប្រូតេអ៊ីនអាចជា សាមញ្ញនិង ស្មុគស្មាញ។ប្រូតេអ៊ីនសាមញ្ញត្រូវបានបង្កើតឡើងពីអាស៊ីតអាមីណូតែប៉ុណ្ណោះខណៈពេលដែល ស្មុគស្មាញប្រូតេអ៊ីន (lipoproteins, chromoproteins, glycoproteins, nucleoproteins ជាដើម) មានប្រូតេអ៊ីន និងផ្នែកដែលមិនមែនជាប្រូតេអ៊ីន។ ក្រូម៉ូសូមប្រូតេអ៊ីនមានផ្នែកដែលមិនមានជាតិប្រូតេអ៊ីន។ ទាំងនេះរួមមាន hemoglobin, myoglobin, chlorophyll, cytochromes ជាដើម។ ដូច្នេះនៅក្នុងសមាសភាពនៃ hemoglobin ខ្សែសង្វាក់ polypeptide នីមួយៗនៃប្រូតេអ៊ីន globin ទាំងបួនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងផ្នែកដែលមិនមែនជាប្រូតេអ៊ីន - heme ដែលនៅចំកណ្តាលមានជាតិដែក។ អ៊ីយ៉ុងដែលផ្តល់ឱ្យអេម៉ូក្លូប៊ីនពណ៌ក្រហម។ ផ្នែកដែលមិនមែនជាប្រូតេអ៊ីន lipoproteinsគឺជា lipid និង glycoproteins- កាបូអ៊ីដ្រាត។ ទាំង lipoproteins និង glycoproteins គឺជាផ្នែកមួយនៃភ្នាសកោសិកា។ នុយក្លេអូប្រូតេអ៊ីនគឺជាស្មុគស្មាញនៃប្រូតេអ៊ីន និងអាស៊ីត nucleic (DNA និង RNA)។ ពួកគេអនុវត្តមុខងារសំខាន់បំផុតនៅក្នុងដំណើរការនៃការផ្ទុក និងការបញ្ជូនព័ត៌មានតំណពូជ។
លក្ខណៈសម្បត្តិប្រូតេអ៊ីន។ប្រូតេអ៊ីនជាច្រើនគឺរលាយក្នុងទឹកបានខ្ពស់ ប៉ុន្តែមានមួយចំនួនក្នុងចំណោមពួកវាដែលរលាយក្នុងដំណោះស្រាយអំបិល អាល់កាឡាំង អាស៊ីត ឬសារធាតុរំលាយសរីរាង្គ។ រចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីន និងសកម្មភាពមុខងាររបស់វាអាស្រ័យលើលក្ខខណ្ឌបរិស្ថាន។ ការបាត់បង់ម៉ូលេគុលប្រូតេអ៊ីននៃរចនាសម្ព័ន្ធរបស់វាខណៈពេលដែលរក្សាបឋមត្រូវបានគេហៅថា ការប្រែពណ៌.

Denaturation កើតឡើងដោយសារតែការផ្លាស់ប្តូរសីតុណ្ហភាព pH សម្ពាធបរិយាកាស ក្រោមឥទ្ធិពលនៃអាស៊ីត អាល់កាឡាំង អំបិលនៃលោហធាតុធ្ងន់ សារធាតុរំលាយសរីរាង្គ។ល។ ដំណើរការបញ្ច្រាសនៃការស្ដាររចនាសម្ព័ន្ធបន្ទាប់បន្សំ និងខ្ពស់ជាងនេះត្រូវបានគេហៅថា ការកែប្រែឡើងវិញទោះយ៉ាងណាក៏ដោយ វាមិនតែងតែអាចទៅរួចនោះទេ។ ការបំបែកពេញលេញនៃម៉ូលេគុលប្រូតេអ៊ីនត្រូវបានគេហៅថា ការបំផ្លិចបំផ្លាញ.
មុខងារប្រូតេអ៊ីន។ប្រូតេអ៊ីនអនុវត្តមុខងារមួយចំនួននៅក្នុងកោសិកា៖ ប្លាស្ទិក (សំណង់) កាតាលីករ (អង់ស៊ីម) ថាមពល សញ្ញា (អ្នកទទួល) កុងតាក់ (ម៉ូទ័រ) ការដឹកជញ្ជូន ការការពារ បទប្បញ្ញត្តិ និងការផ្ទុក។
មុខងារបង្កើតប្រូតេអ៊ីនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងវត្តមានរបស់វានៅក្នុងភ្នាសកោសិកា និងសមាសធាតុរចនាសម្ព័ន្ធនៃកោសិកា។ ថាមពល - ដោយសារតែការពិតដែលថាក្នុងអំឡុងពេលបំបែកប្រូតេអ៊ីន 1 ក្រាមថាមពល 17.2 kJ ត្រូវបានបញ្ចេញ។ ប្រូតេអ៊ីន receptor Membrane ត្រូវបានចូលរួមយ៉ាងសកម្មក្នុងការយល់ឃើញនៃសញ្ញាបរិស្ថាន និងការបញ្ជូនរបស់ពួកគេតាមរយៈកោសិកា ក៏ដូចជានៅក្នុងការទទួលស្គាល់អន្តរកោសិកាផងដែរ។ បើគ្មានប្រូតេអ៊ីន ចលនានៃកោសិកា និងសារពាង្គកាយទាំងមូលគឺមិនអាចទៅរួចទេព្រោះវាបង្កើតជាមូលដ្ឋាននៃ flagella និង cilia ហើយក៏ផ្តល់នូវការកន្ត្រាក់សាច់ដុំ និងចលនានៃសមាសធាតុ intracellular ផងដែរ។ នៅក្នុងឈាមរបស់មនុស្ស និងសត្វជាច្រើន ប្រូតេអ៊ីនអេម៉ូក្លូប៊ីនផ្ទុកអុកស៊ីសែន និងផ្នែកនៃកាបូនឌីអុកស៊ីត ខណៈដែលប្រូតេអ៊ីនផ្សេងទៀតដឹកជញ្ជូនអ៊ីយ៉ុង និងអេឡិចត្រុង។ តួនាទីការពាររបស់ប្រូតេអ៊ីនត្រូវបានផ្សារភ្ជាប់ជាចម្បងជាមួយនឹងភាពស៊ាំ ចាប់តាំងពីប្រូតេអ៊ីន interferon អាចបំផ្លាញមេរោគជាច្រើន ហើយប្រូតេអ៊ីនអង្គបដិប្រាណរារាំងការវិវត្តនៃបាក់តេរី និងភ្នាក់ងារបរទេសផ្សេងទៀត។ មានអរម៉ូនជាច្រើនក្នុងចំណោមប្រូតេអ៊ីន និង peptides ឧទាហរណ៍ អាំងស៊ុយលីនអរម៉ូនលំពែង ដែលគ្រប់គ្រងកំហាប់គ្លុយកូសក្នុងឈាម។ នៅក្នុងសារពាង្គកាយមួយចំនួន ប្រូតេអ៊ីនអាចត្រូវបានរក្សាទុកក្នុងបំរុង ដូចជានៅក្នុងគ្រាប់ធញ្ញជាតិ ឬប្រូតេអ៊ីននៃស៊ុតមាន់។
អាស៊ីតនុយក្លេអ៊ីក
អាស៊ីតនុយក្លេអ៊ីកគឺជាជីវប៉ូលីម័រដែលម៉ូណូមឺរគឺជានុយក្លេអូទីត។ បច្ចុប្បន្ននេះអាស៊ីតនុយក្លេអ៊ីកពីរប្រភេទត្រូវបានគេស្គាល់៖ ribonucleic (RNA) និង deoxyribonucleic (DNA) ។
នុយក្លេអូទីតបង្កើតឡើងដោយមូលដ្ឋានអាសូត សំណល់ស្ករ pentose និងសំណល់អាស៊ីតផូស្វ័រ។ លក្ខណៈពិសេសនៃនុយក្លេអូទីតត្រូវបានកំណត់ជាចម្បងដោយមូលដ្ឋានអាសូតដែលបង្កើតសមាសភាពរបស់វា ដូច្នេះសូម្បីតែតាមលក្ខខណ្ឌ នុយក្លេអូទីតត្រូវបានកំណត់ដោយអក្សរដំបូងនៃឈ្មោះរបស់វា។ សមាសធាតុនៃនុយក្លេអូទីតអាចរួមបញ្ចូលមូលដ្ឋានអាសូតចំនួនប្រាំ៖ អាឌីនីន (A), ហ្គានីន (G), ធីមីន (T), យូរ៉ាស៊ីល (យូ) និងស៊ីតូស៊ីន (ស៊ី) ។ pentoses នៃ nucleotides - ribose និង deoxyribose - កំណត់ថាតើ nucleotide មួយណានឹងត្រូវបានបង្កើតឡើង - ribonucleotide ឬ deoxyribonucleotide ។ Ribonucleotides គឺជា RNA monomers ពួកគេអាចដើរតួជាម៉ូលេគុលសញ្ញា (cAMP) និងជាផ្នែកមួយនៃសមាសធាតុថាមពលខ្ពស់ដូចជា ATP និង coenzymes ដូចជា NADP, NAD, FAD ជាដើម ហើយ deoxyribonucleotides គឺជាផ្នែកមួយនៃ DNA ។

អាស៊ីត deoxyribonucleic (DNA)- ជីវប៉ូលីម័រដែលមានខ្សែទ្វេ ដែលជាម៉ូណូមឺរដែលជា deoxyribonucleotides ។ សមាសភាពនៃ deoxyribonucleotides រួមមានមូលដ្ឋានអាសូតចំនួន 4 ក្នុងចំណោម 5 ដែលអាចធ្វើទៅបាន - adenine (A), thymine (T), guanine (G) ឬ cytosine (C) ក៏ដូចជា deoxyribose និងសំណល់អាស៊ីតផូស្វ័រ។ នុយក្លេអូទីតនៅក្នុងខ្សែសង្វាក់ DNA ត្រូវបានភ្ជាប់គ្នាទៅវិញទៅមកតាមរយៈសំណល់អាស៊ីត orthophosphoric បង្កើតជាចំណង phosphodiester ។ នៅពេលដែលម៉ូលេគុលដែលមានខ្សែពីរត្រូវបានបង្កើតឡើង មូលដ្ឋានអាសូតត្រូវបានដឹកនាំទៅខាងក្នុងនៃម៉ូលេគុល។ ទោះជាយ៉ាងណាក៏ដោយការភ្ជាប់ខ្សែសង្វាក់ DNA មិនកើតឡើងដោយចៃដន្យទេ - មូលដ្ឋានអាសូតនៃខ្សែសង្វាក់ផ្សេងៗគ្នាត្រូវបានភ្ជាប់គ្នាទៅវិញទៅមកដោយចំណងអ៊ីដ្រូសែនយោងទៅតាមគោលការណ៍នៃការបំពេញបន្ថែម: adenine ត្រូវបានភ្ជាប់ទៅនឹង thymine ដោយចំណងអ៊ីដ្រូសែនពីរ (A \u003d T) និង guanine និង cytosine ។ ដោយបី (G $ ≡ $ C) ។

សម្រាប់នាងត្រូវបានកំណត់ ច្បាប់ស្តីពីការចោទប្រកាន់:
- ចំនួននុយក្លេអូទីត DNA ដែលមានអាឌីនីន គឺស្មើនឹងចំនួននុយក្លេអូទីតដែលមាន thymine (A=T)។
- ចំនួននុយក្លេអូទីត DNA ដែលមាន guanine គឺស្មើនឹងចំនួននុយក្លេអូទីតដែលមានស៊ីតូស៊ីន (G$≡$C)។
- ផលបូកនៃ deoxyribonucleotides ដែលមាន adenine និង guanine គឺស្មើនឹងផលបូកនៃ deoxyribonucleotides ដែលមាន thymine និង cytosine (A+G = T+C) ។
- សមាមាត្រនៃផលបូកនៃ deoxyribonucleotides ដែលមាន adenine និង thymine ទៅនឹងផលបូក deoxyribonucleotides ដែលមាន guanine និង cytosine អាស្រ័យលើប្រភេទនៃសារពាង្គកាយ។
រចនាសម្ព័ន DNA ត្រូវបានបកស្រាយដោយ F. Crick និង D. Watson (រង្វាន់ណូបែលផ្នែកសរីរវិទ្យា ឬវេជ្ជសាស្ត្រ ឆ្នាំ 1962)។ យោងតាមគំរូរបស់ពួកគេ ម៉ូលេគុល DNA គឺជា helix ដៃស្តាំពីរ។ ចម្ងាយរវាងនុយក្លេអូទីតនៅក្នុងខ្សែ DNA គឺ 0.34 nm ។

ទ្រព្យសម្បត្តិសំខាន់បំផុតរបស់ DNA គឺសមត្ថភាពក្នុងការចម្លង (ការចម្លងដោយខ្លួនឯង) ។ មុខងារសំខាន់នៃ DNA គឺការផ្ទុក និងការបញ្ជូនព័ត៌មានតំណពូជ ដែលត្រូវបានសរសេរក្នុងទម្រង់នៃលំដាប់នុយក្លេអូទីត។ ស្ថេរភាពនៃម៉ូលេគុល DNA ត្រូវបានរក្សាដោយប្រព័ន្ធជួសជុលដ៏មានអានុភាព (ការស្តារឡើងវិញ) ប៉ុន្តែសូម្បីតែពួកគេក៏មិនអាចលុបបំបាត់ផលប៉ះពាល់អវិជ្ជមានបានទាំងស្រុងដែរ ដែលនៅទីបំផុតនាំទៅរកការផ្លាស់ប្តូរ។ DNA នៃកោសិកា eukaryotic ត្រូវបានប្រមូលផ្តុំនៅក្នុង nucleus, mitochondria និង plastids ខណៈពេលដែលកោសិកា prokaryotic មានទីតាំងនៅដោយផ្ទាល់នៅក្នុង cytoplasm ។ DNA នុយក្លេអ៊ែរគឺជាមូលដ្ឋាននៃក្រូម៉ូសូម វាត្រូវបានតំណាងដោយម៉ូលេគុលបើកចំហ។ DNA នៃ mitochondria, plastids និង prokaryotes មានរាងជារង្វង់។
អាស៊ីត Ribonucleic (RNA)- biopolymer ដែល monomers គឺ ribonucleotides ។ ពួកគេក៏មានមូលដ្ឋានអាសូតចំនួនបួនផងដែរ - adenine (A), uracil (U), guanine (G) ឬ cytosine (C) ដោយហេតុនេះខុសគ្នាពី DNA នៅក្នុងមូលដ្ឋានមួយ (ជំនួសឱ្យ thymine, RNA មានផ្ទុក uracil) ។ សំណល់ជាតិស្ករ pentose នៅក្នុង ribonucleotides ត្រូវបានតំណាងដោយ ribose ។ RNA ភាគច្រើនជាម៉ូលេគុលដែលមានខ្សែតែមួយ លើកលែងតែមេរោគមួយចំនួន។ RNA មានបីប្រភេទសំខាន់ៗ៖ ព័ត៌មាន ឬគំរូ (mRNA, mRNA), ribosomal (rRNA) និងការដឹកជញ្ជូន (tRNA) ។ ពួកគេទាំងអស់ត្រូវបានបង្កើតឡើងនៅក្នុងដំណើរការ ប្រតិចារិក- សរសេរឡើងវិញពីម៉ូលេគុល DNA ។
និង RNAs បង្កើតជាប្រភាគតូចបំផុតនៃ RNA នៅក្នុងកោសិកាមួយ (2-4%) ដែលត្រូវបានទូទាត់ដោយភាពចម្រុះរបស់វា ដោយសារកោសិកាមួយអាចផ្ទុក mRNA ខុសៗគ្នារាប់ពាន់។ ទាំងនេះគឺជាម៉ូលេគុលខ្សែតែមួយ ដែលជាគំរូសម្រាប់ការសំយោគខ្សែសង្វាក់ polypeptide ។ ព័ត៌មានអំពីរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីនត្រូវបានកត់ត្រានៅក្នុងពួកវាក្នុងទម្រង់ជាលំដាប់នៃនុយក្លេអូទីត ហើយអាស៊ីតអាមីណូនីមួយៗបញ្ចូលកូដនុយក្លេអូទីតបីដង - កូឌុន.
រ RNA គឺជាប្រភេទ RNA ច្រើនបំផុតនៅក្នុងកោសិកា (រហូតដល់ 80%) ។ ទម្ងន់ម៉ូលេគុលរបស់ពួកគេជាមធ្យម 3000-5000; ត្រូវបានបង្កើតឡើងនៅក្នុង nucleoli និងជាផ្នែកមួយនៃសរីរាង្គកោសិកា - ribosomes ។ rRNAs ក៏ហាក់ដូចជាដើរតួក្នុងការសំយោគប្រូតេអ៊ីនផងដែរ។
ធ RNA គឺជាម៉ូលេគុល RNA តូចបំផុតព្រោះវាមាននុយក្លេអូទីតតែ 73-85 ប៉ុណ្ណោះ។ ចំណែករបស់ពួកគេនៃចំនួនសរុបនៃ RNA កោសិកាគឺប្រហែល 16% ។ មុខងាររបស់ tRNA គឺជាការដឹកជញ្ជូនអាស៊ីតអាមីណូទៅកាន់កន្លែងសំយោគប្រូតេអ៊ីន (នៅលើ ribosomes)។ រូបរាងនៃម៉ូលេគុល tRNA ប្រហាក់ប្រហែលនឹងស្លឹក clover ។ នៅចុងម្ខាងនៃម៉ូលេគុលមានកន្លែងសម្រាប់ភ្ជាប់អាស៊ីតអាមីណូ ហើយនៅក្នុងរង្វិលជុំមួយមាននុយក្លេអូទីតបីដែលបំពេញបន្ថែមជាមួយ mRNA codon និងកំណត់ថាតើអាស៊ីតអាមីណូមួយណាដែល tRNA នឹងផ្ទុក - ថ្នាំ anticodon ។

គ្រប់ប្រភេទនៃ RNA ចូលរួមយ៉ាងសកម្មក្នុងការអនុវត្តព័ត៌មានតំណពូជ ដែលត្រូវបានសរសេរឡើងវិញពី DNA ទៅ mRNA ហើយការសំយោគប្រូតេអ៊ីនចុងក្រោយត្រូវបានអនុវត្ត។ tRNA នៅក្នុងដំណើរការនៃការសំយោគប្រូតេអ៊ីនផ្តល់នូវអាស៊ីតអាមីណូទៅកាន់ ribosomes ហើយ rRNA គឺជាផ្នែកមួយនៃ ribosomes ដោយផ្ទាល់។
អាស៊ីត Adenosine triphosphoric (ATP)គឺជានុយក្លេអូទីតដែលមាន បន្ថែមពីលើមូលដ្ឋានអាសូតនៃអាឌីនីន និងសំណល់រីបូស សំណល់អាស៊ីតផូស្វ័របី។ ចំណងរវាងសំណល់ផូស្វ័រពីរចុងក្រោយគឺ macroergic (42 kJ / mol នៃថាមពលត្រូវបានបញ្ចេញកំឡុងពេលបំបែក) ខណៈពេលដែលចំណងគីមីស្តង់ដារក្នុងអំឡុងពេលពុះផ្តល់ 12 kJ / mol ។ ប្រសិនបើត្រូវការថាមពល ចំណង macroergic នៃ ATP ត្រូវបានបំបែក អាស៊ីត adenosine diphosphoric (ADP) សំណល់ផូស្វ័រត្រូវបានបង្កើតឡើង ហើយថាមពលត្រូវបានបញ្ចេញ៖
ATP + H 2 O $ → $ ADP + H 3 PO 4 + 42 kJ ។
ADP ក៏អាចត្រូវបានបំបែកទៅជា AMP (អាស៊ីត adenosine monophosphoric) និងសំណល់អាស៊ីតផូស្វ័រ៖
ADP + H 2 O $ → $ AMP + H 3 PO 4 + 42 kJ ។
នៅក្នុងដំណើរការនៃការរំលាយអាហារថាមពល (កំឡុងពេលដកដង្ហើម ការ fermentation) ក៏ដូចជានៅក្នុងដំណើរការនៃការធ្វើរស្មីសំយោគ ADP ភ្ជាប់សំណល់ផូស្វ័រ ហើយប្រែទៅជា ATP ។ ប្រតិកម្មនៃការងើបឡើងវិញ ATP ត្រូវបានគេហៅថា ផូស្វ័រ. ATP គឺជាប្រភពថាមពលសកលសម្រាប់ដំណើរការជីវិតទាំងអស់នៃសារពាង្គកាយមានជីវិត។
ការសិក្សាអំពីសមាសធាតុគីមីនៃកោសិកានៃសារពាង្គកាយមានជីវិតទាំងអស់បានបង្ហាញថាពួកវាមានធាតុគីមីដូចគ្នា សារធាតុគីមីដែលបំពេញមុខងារដូចគ្នា។ លើសពីនេះទៅទៀត បំណែកនៃ DNA ដែលផ្ទេរពីសារពាង្គកាយមួយទៅសារពាង្គកាយមួយទៀតនឹងដំណើរការនៅក្នុងវា ហើយប្រូតេអ៊ីនដែលសំយោគដោយបាក់តេរី ឬផ្សិតនឹងដើរតួជាអរម៉ូន ឬអង់ស៊ីមនៅក្នុងខ្លួនមនុស្ស។ នេះគឺជាភស្ដុតាងមួយនៃការរួបរួមនៃប្រភពដើមនៃពិភពសរីរាង្គ។

រចនាសម្ព័ន្ធកោសិកា។ ទំនាក់ទំនងនៃរចនាសម្ព័ន្ធ និងមុខងារនៃផ្នែក និងសរីរាង្គនៃកោសិកា គឺជាមូលដ្ឋាននៃភាពសុចរិតរបស់វា
រចនាសម្ព័ន្ធកោសិកា
រចនាសម្ព័ន្ធនៃកោសិកា prokaryotic និង eukaryotic
សមាសធាតុរចនាសម្ព័ន្ធសំខាន់ៗនៃកោសិកាគឺភ្នាសប្លាស្មា ស៊ីតូប្លាស និងបរិធានតំណពូជ។ អាស្រ័យលើលក្ខណៈរបស់អង្គការ កោសិកាពីរប្រភេទសំខាន់ៗត្រូវបានសម្គាល់៖ prokaryotic និង eukaryotic ។ ភាពខុសគ្នាសំខាន់រវាងកោសិកា prokaryotic និង eukaryotic គឺជាការរៀបចំឧបករណ៍តំណពូជរបស់ពួកគេ៖ នៅក្នុង prokaryotes វាមានទីតាំងនៅដោយផ្ទាល់នៅក្នុង cytoplasm (តំបន់នៃ cytoplasm ត្រូវបានគេហៅថា នុយក្លីអ៊ីត) ហើយមិនត្រូវបានបំបែកចេញពីវាដោយរចនាសម្ព័ន្ធភ្នាសទេ ខណៈពេលដែលនៅក្នុង eukaryotes ភាគច្រើននៃ DNA ត្រូវបានប្រមូលផ្តុំនៅក្នុងស្នូលដែលហ៊ុំព័ទ្ធដោយភ្នាសទ្វេ។ លើសពីនេះទៀតព័ត៌មានហ្សែននៃកោសិកា prokaryotic ដែលមានទីតាំងនៅ nucleoid ត្រូវបានកត់ត្រានៅក្នុងម៉ូលេគុល DNA រាងជារង្វង់ខណៈពេលដែលនៅក្នុង eukaryotes ម៉ូលេគុល DNA មិនត្រូវបានបិទ។
មិនដូច eukaryotes ទេ cytoplasm នៃកោសិកា prokaryotic ក៏មានបរិមាណសរីរាង្គតិចតួចផងដែរ ខណៈដែលកោសិកា eukaryotic ត្រូវបានកំណត់លក្ខណៈដោយភាពខុសគ្នាសំខាន់ៗនៃរចនាសម្ព័ន្ធទាំងនេះ។
រចនាសម្ព័ន្ធនិងមុខងារនៃភ្នាសជីវសាស្រ្ត
រចនាសម្ព័ន្ធនៃ biomembrane ។ភ្នាសកោសិកា និងសរីរាង្គភ្នាសនៃកោសិកា eukaryotic ចែករំលែកសមាសភាពគីមីទូទៅ និងរចនាសម្ព័ន្ធ។ ពួកវារួមមាន lipid ប្រូតេអ៊ីន និងកាបូអ៊ីដ្រាត។ ភ្នាសរំអិលត្រូវបានតំណាងជាចម្បងដោយ phospholipids និង cholesterol ។ ប្រូតេអ៊ីនភ្នាសភាគច្រើនគឺជាប្រូតេអ៊ីនស្មុគស្មាញដូចជា glycoproteins ។ កាបូអ៊ីដ្រាតមិនកើតឡើងដោយខ្លួនឯងនៅក្នុងភ្នាសទេពួកគេត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងប្រូតេអ៊ីននិងខ្លាញ់។ កម្រាស់នៃភ្នាសគឺ 7-10 nm ។

យោងតាមគំរូ mosaic សារធាតុរាវដែលទទួលយកនាពេលបច្ចុប្បន្ននៃរចនាសម្ព័ន្ធភ្នាស lipid បង្កើតជាស្រទាប់ទ្វេរឬ lipid bilayerដែលក្នុងនោះ "ក្បាល" hydrophilic នៃម៉ូលេគុល lipid ត្រូវបានប្រែទៅជាខាងក្រៅហើយ "កន្ទុយ" hydrophobic ត្រូវបានលាក់នៅខាងក្នុងភ្នាស។ "កន្ទុយ" ទាំងនេះដោយសារតែ hydrophobicity របស់ពួកគេធានានូវការបំបែកនៃដំណាក់កាល aqueous នៃបរិយាកាសខាងក្នុងនៃកោសិកានិងបរិស្ថានរបស់វា។ ប្រូតេអ៊ីនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹង lipid តាមរយៈប្រភេទផ្សេងៗនៃអន្តរកម្ម។ ប្រូតេអ៊ីនមួយចំនួនមានទីតាំងនៅលើផ្ទៃនៃភ្នាស។ ប្រូតេអ៊ីនបែបនេះត្រូវបានគេហៅថា គ្រឿងកុំព្យូទ័រ, ឬ លើផ្ទៃ. ប្រូតេអ៊ីនផ្សេងទៀតត្រូវបានជ្រមុជដោយផ្នែកឬទាំងស្រុងនៅក្នុងភ្នាស - ទាំងនេះគឺ អាំងតេក្រាលឬ ប្រូតេអ៊ីនលិចទឹក។. ប្រូតេអ៊ីន Membrane អនុវត្តរចនាសម្ព័ន្ធ ការដឹកជញ្ជូន កាតាលីករ អ្នកទទួល និងមុខងារផ្សេងៗទៀត។
Membranes មិនដូចគ្រីស្តាល់ទេសមាសធាតុរបស់វាមានចលនាឥតឈប់ឈរដែលជាលទ្ធផលនៃចន្លោះប្រហោងលេចឡើងរវាងម៉ូលេគុលខ្លាញ់ - រន្ធញើសដែលសារធាតុផ្សេងៗអាចចូលឬចេញពីកោសិកា។
ភ្នាសជីវសាស្រ្តមានភាពខុសប្លែកគ្នានៅក្នុងទីតាំងរបស់ពួកគេនៅក្នុងកោសិកា សមាសធាតុគីមី និងមុខងាររបស់វា។ ប្រភេទសំខាន់នៃភ្នាសគឺប្លាស្មានិងខាងក្នុង។ ភ្នាសប្លាស្មាមានផ្ទុកជាតិខ្លាញ់ប្រហែល 45% (រួមទាំង glycolipids) ប្រូតេអ៊ីន 50% និងកាបូអ៊ីដ្រាត 5% ។ ខ្សែសង្វាក់នៃកាបូអ៊ីដ្រាតដែលបង្កើតជាប្រូតេអ៊ីនស្មុគស្មាញ - glycoproteins និង lipids-glycolipids ស្មុគស្មាញលាតសន្ធឹងពីលើផ្ទៃនៃភ្នាស។ plasmalemmal glycoproteins គឺជាក់លាក់បំផុត។ ដូច្នេះជាឧទាហរណ៍តាមរយៈពួកវាមានការទទួលស្គាល់គ្នាទៅវិញទៅមកនៃកោសិការួមទាំងមេជីវិតឈ្មោលនិងស៊ុត។
នៅលើផ្ទៃនៃកោសិកាសត្វ ខ្សែសង្វាក់កាបូអ៊ីដ្រាតបង្កើតជាស្រទាប់ផ្ទៃស្តើង - glycocalyx ។វាត្រូវបានគេរកឃើញនៅក្នុងកោសិកាសត្វស្ទើរតែទាំងអស់ ប៉ុន្តែភាពធ្ងន់ធ្ងររបស់វាមិនដូចគ្នាទេ (10-50 microns)។ glycocalyx ផ្តល់នូវការតភ្ជាប់ដោយផ្ទាល់នៃកោសិកាជាមួយបរិយាកាសខាងក្រៅ ការរំលាយអាហារក្រៅកោសិកាកើតឡើងនៅក្នុងវា; អ្នកទទួលមានទីតាំងនៅ glycocalyx ។ កោសិកានៃបាក់តេរី រុក្ខជាតិ និងផ្សិត បន្ថែមពីលើប្លាស្មាម៉ាម៉ា ក៏ត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសកោសិកាផងដែរ។
ភ្នាសខាងក្នុងកោសិកា eukaryotic កំណត់ផ្នែកផ្សេងៗនៃកោសិកា បង្កើតជាប្រភេទនៃ "compartments" - បន្ទប់ដែលរួមចំណែកដល់ការបំបែកដំណើរការផ្សេងៗនៃការរំលាយអាហារ និងថាមពល។ ពួកវាអាចមានភាពខុសប្លែកគ្នានៅក្នុងសមាសភាព និងមុខងារគីមី ប៉ុន្តែពួកគេរក្សានូវផែនការទូទៅនៃរចនាសម្ព័ន្ធ។
មុខងារភ្នាស៖
- ការកំណត់។វាមាននៅក្នុងការពិតដែលថាពួកវាបំបែកចន្លោះខាងក្នុងនៃកោសិកាពីបរិយាកាសខាងក្រៅ។ ភ្នាសគឺអាចជ្រាបចូលបានពាក់កណ្តាល ពោលគឺមានតែសារធាតុទាំងនោះដែលចាំបាច់សម្រាប់កោសិកាអាចយកឈ្នះវាបានដោយសេរី ខណៈពេលដែលមានយន្តការសម្រាប់ដឹកជញ្ជូនសារធាតុចាំបាច់។
- អ្នកទទួល។វាត្រូវបានភ្ជាប់ជាចម្បងជាមួយនឹងការយល់ឃើញនៃសញ្ញាបរិស្ថាន និងការផ្ទេរព័ត៌មាននេះទៅក្នុងក្រឡា។ ប្រូតេអ៊ីនទទួលពិសេសទទួលខុសត្រូវចំពោះមុខងារនេះ។ ប្រូតេអ៊ីន Membrane ក៏ទទួលខុសត្រូវចំពោះការទទួលស្គាល់កោសិកាយោងទៅតាមគោលការណ៍ "មិត្តឬសត្រូវ" ក៏ដូចជាសម្រាប់ការបង្កើតទំនាក់ទំនងរវាងកោសិកាដែលត្រូវបានសិក្សាច្រើនបំផុតគឺ synapses នៃកោសិកាសរសៃប្រសាទ។
- កាតាលីករ។ស្មុគស្មាញអង់ស៊ីមជាច្រើនមានទីតាំងនៅលើភ្នាសដែលជាលទ្ធផលនៃដំណើរការសំយោគដែលពឹងផ្អែកខ្លាំងកើតឡើងលើពួកវា។
- ការបំប្លែងថាមពល។ទាក់ទងនឹងការបង្កើតថាមពល ការផ្ទុករបស់វាក្នុងទម្រង់ ATP និងការចំណាយ។
- ការបែងចែកផ្នែក។ភ្នាសក៏កំណត់ចន្លោះខាងក្នុងកោសិកា ដោយហេតុនេះបំបែកសារធាតុដំបូងនៃប្រតិកម្ម និងអង់ស៊ីមដែលអាចអនុវត្តប្រតិកម្មដែលត្រូវគ្នា។
- ការបង្កើតទំនាក់ទំនងអន្តរកោសិកា។ទោះបីជាការពិតដែលថាកម្រាស់នៃភ្នាសគឺតូចដូច្នេះវាមិនអាចសម្គាល់បានដោយភ្នែកទទេក៏ដោយ ក៏វាដើរតួជារបាំងដែលអាចទុកចិត្តបានសម្រាប់អ៊ីយ៉ុង និងម៉ូលេគុល ជាពិសេសសារធាតុរលាយក្នុងទឹក ហើយម្យ៉ាងវិញទៀត វាធានាការផ្ទេររបស់ពួកគេទៅក្នុងក្រឡា និងចេញ។
- ការដឹកជញ្ជូន។

ការដឹកជញ្ជូនភ្នាស។ដោយសារតែកោសិកាដែលជាប្រព័ន្ធជីវសាស្រ្តបឋមគឺជាប្រព័ន្ធបើកចំហដើម្បីធានាបាននូវការរំលាយអាហារនិងថាមពលរក្សា homeostasis ការលូតលាស់ការឆាប់ខឹងនិងដំណើរការផ្សេងទៀតការផ្ទេរសារធាតុតាមរយៈភ្នាសត្រូវបានទាមទារ - ការដឹកជញ្ជូនភ្នាស។ បច្ចុប្បន្ននេះការដឹកជញ្ជូនសារធាតុឆ្លងកាត់ភ្នាសកោសិកាត្រូវបានបែងចែកទៅជាសកម្មអកម្ម endo- និង exocytosis ។
ការដឹកជញ្ជូនអកម្មគឺជាប្រភេទនៃការដឹកជញ្ជូនដែលកើតឡើងដោយមិនចំណាយថាមពលពីកំហាប់ខ្ពស់ទៅទាប។ ម៉ូលេគុលតូចៗដែលមិនរលាយក្នុងប៉ូល (O 2, CO 2) ងាយជ្រាបចូលទៅក្នុងកោសិកាដោយ ការសាយភាយសាមញ្ញ. មិនរលាយក្នុង lipid រួមទាំងភាគល្អិតតូចៗដែលត្រូវបានចោទប្រកាន់ត្រូវបានប្រមូលដោយប្រូតេអ៊ីនក្រុមហ៊ុនដឹកជញ្ជូនឬឆ្លងកាត់បណ្តាញពិសេស (គ្លុយកូស, អាស៊ីតអាមីណូ, K +, PO 4 3-) ។ ប្រភេទនៃការដឹកជញ្ជូនអកម្មនេះត្រូវបានគេហៅថា សម្រួលដល់ការសាយភាយ. ទឹកចូលទៅក្នុងកោសិកាតាមរយៈរន្ធញើសក្នុងដំណាក់កាល lipid ក៏ដូចជាតាមរយៈបណ្តាញពិសេសដែលតម្រង់ជួរជាមួយប្រូតេអ៊ីន។ ការដឹកជញ្ជូនទឹកឆ្លងកាត់ភ្នាសត្រូវបានគេហៅថា osmosis ។

Osmosis គឺមានសារៈសំខាន់ខ្លាំងណាស់ក្នុងជីវិតរបស់កោសិកា ព្រោះប្រសិនបើវាត្រូវបានដាក់ក្នុងដំណោះស្រាយដែលមានកំហាប់អំបិលខ្ពស់ជាងនៅក្នុងដំណោះស្រាយកោសិកា នោះទឹកនឹងចាប់ផ្តើមចេញពីកោសិកា ហើយបរិមាណនៃសារធាតុរស់នៅនឹងចាប់ផ្តើមថយចុះ។ . នៅក្នុងកោសិកាសត្វ កោសិកាទាំងមូលរួមតូច ហើយនៅក្នុងកោសិការុក្ខជាតិ cytoplasm យឺតនៅពីក្រោយជញ្ជាំងកោសិកា ដែលត្រូវបានគេហៅថា plasmolysis. នៅពេលដែលកោសិកាមួយត្រូវបានដាក់ក្នុងដំណោះស្រាយដែលមានកំហាប់តិចជាង cytoplasm នោះទឹកត្រូវបានដឹកជញ្ជូនក្នុងទិសដៅផ្ទុយ - ចូលទៅក្នុងកោសិកា។ ទោះយ៉ាងណាក៏ដោយ មានដែនកំណត់ចំពោះការពង្រីកនៃភ្នាស cytoplasmic ហើយកោសិកាសត្វនឹងរហែកជាយថាហេតុ ខណៈពេលដែលនៅក្នុងកោសិការុក្ខជាតិ វាមិនត្រូវបានអនុញ្ញាតដោយជញ្ជាំងកោសិកាដ៏រឹងមាំនោះទេ។ បាតុភូតនៃការបំពេញចន្លោះខាងក្នុងទាំងមូលនៃកោសិកាជាមួយនឹងមាតិកាកោសិកាត្រូវបានគេហៅថា deplasmolysis. កំហាប់អំបិលក្នុងកោសិកាគួរតែត្រូវយកមកពិចារណានៅពេលរៀបចំថ្នាំ ជាពិសេសសម្រាប់ការគ្រប់គ្រងតាមសរសៃឈាម ព្រោះនេះអាចនាំឱ្យខូចកោសិកាឈាម (សម្រាប់បញ្ហានេះ ដំណោះស្រាយអំបិលដែលមានកំហាប់សូដ្យូមក្លរួ ០,៩% ត្រូវបានប្រើ)។ នេះមិនសំខាន់តិចទេក្នុងការដាំដុះកោសិកា និងជាលិកា ក៏ដូចជាសរីរាង្គរបស់សត្វ និងរុក្ខជាតិ។

ការដឹកជញ្ជូនសកម្មដំណើរការជាមួយនឹងការចំណាយថាមពល ATP ពីកំហាប់ទាបនៃសារធាតុមួយទៅកម្រិតខ្ពស់។ វាត្រូវបានអនុវត្តដោយជំនួយពីប្រូតេអ៊ីនពិសេស - ស្នប់។ ប្រូតេអ៊ីនបូមអ៊ីយ៉ុង K +, Na +, Ca 2+ និងផ្សេងទៀតតាមរយៈភ្នាសដែលរួមចំណែកដល់ការដឹកជញ្ជូនសារធាតុសរីរាង្គសំខាន់ៗក៏ដូចជាការលេចឡើងនៃសរសៃប្រសាទ។ល។
ជំងឺ endocytosis- នេះគឺជាដំណើរការសកម្មនៃការស្រូបយកសារធាតុដោយកោសិកា ដែលភ្នាសបង្កើតជា invagination ហើយបន្ទាប់មកបង្កើតជាភ្នាសភ្នាស។ phagosomesដែលមានវត្ថុស្រូប។ lysosome បឋមបន្ទាប់មកប្រសព្វជាមួយនឹង phagosome ដើម្បីបង្កើត lysosome ទីពីរ, ឬ phagolysosome, ឬ vacuole រំលាយអាហារ. ខ្លឹមសារនៃ vesicle ត្រូវបានបំបែកដោយអង់ស៊ីម lysosome ហើយផលិតផលបំបែកត្រូវបានស្រូបយក និង assimilated ដោយកោសិកា។ សំណល់ដែលមិនបានរំលាយត្រូវបានយកចេញពីកោសិកាដោយ exocytosis ។ ជំងឺ endocytosis មានពីរប្រភេទសំខាន់ៗគឺ phagocytosis និង pinocytosis ។
ជំងឺ Phagocytosisគឺជាដំណើរការនៃការចាប់យកដោយផ្ទៃក្រឡា និងការស្រូបយកភាគល្អិតរឹងដោយកោសិកា និង pinocytosis- វត្ថុរាវ។ Phagocytosis កើតឡើងជាចម្បងនៅក្នុងកោសិកាសត្វ (សត្វកោសិកាតែមួយ leukocytes មនុស្ស) វាផ្តល់នូវអាហាររូបត្ថម្ភរបស់ពួកគេហើយជារឿយៗការពាររាងកាយ។ តាមវិធីនៃ pinocytosis ការស្រូបយកប្រូតេអ៊ីន អង់ទីហ្សែន-អង់ទីគ័រស្មុគ្រស្មាញក្នុងដំណើរការនៃប្រតិកម្មនៃប្រព័ន្ធភាពស៊ាំកើតឡើង។ ទោះជាយ៉ាងណាក៏ដោយ មេរោគជាច្រើនក៏ចូលទៅក្នុងកោសិកាដោយវិធីនៃ pinocytosis ឬ phagocytosis ផងដែរ។ នៅក្នុងកោសិកានៃរុក្ខជាតិ និងផ្សិត phagocytosis គឺមិនអាចអនុវត្តបានទេព្រោះវាត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសកោសិកាដ៏រឹងមាំ។

Exocytosisគឺជាដំណើរការបញ្ច្រាសនៃជំងឺ endocytosis ។ ដូច្នេះ សំណល់អាហារដែលមិនបានរំលាយត្រូវបានបញ្ចេញចេញពីបំពង់រំលាយអាហារ សារធាតុចាំបាច់សម្រាប់ជីវិតរបស់កោសិកា និងសារពាង្គកាយទាំងមូលត្រូវបានដកចេញ។ ឧទាហរណ៍ ការបញ្ជូនសរសៃប្រសាទកើតឡើងដោយសារតែការបញ្ចេញសារជាតិគីមីដោយណឺរ៉ូនដែលបញ្ជូនកម្លាំងជំរុញ - អ្នកសម្របសម្រួលហើយនៅក្នុងកោសិការុក្ខជាតិ កាបូអ៊ីដ្រាតជំនួយនៃភ្នាសកោសិកាត្រូវបានបញ្ចេញតាមរបៀបនេះ។
ជញ្ជាំងកោសិកានៃកោសិការុក្ខជាតិ ផ្សិត និងបាក់តេរី។នៅខាងក្រៅភ្នាសកោសិកាអាចលាក់បាំងនូវក្របខ័ណ្ឌដ៏រឹងមាំមួយ - ភ្នាសកោសិកា,ឬ ជញ្ជាំងកោសិកា។
នៅក្នុងរុក្ខជាតិ ជញ្ជាំងកោសិកាត្រូវបានបង្កើតឡើង សែលុយឡូសខ្ចប់ជាបាច់នៃ 50-100 ម៉ូលេគុល។ ចន្លោះរវាងពួកវាត្រូវបានបំពេញដោយទឹកនិងកាបូអ៊ីដ្រាតផ្សេងទៀត។ ភ្នាសកោសិការុក្ខជាតិត្រូវបានទម្លុះដោយ tubules - plasmodesmataតាមរយៈការដែលភ្នាសនៃ reticulum endoplasmic ឆ្លងកាត់។ plasmodesmata ដឹកជញ្ជូនសារធាតុរវាងកោសិកា។ ទោះជាយ៉ាងណាក៏ដោយ ការដឹកជញ្ជូនសារធាតុដូចជាទឹកក៏អាចកើតឡើងតាមជញ្ជាំងកោសិកាខ្លួនឯងដែរ។ យូរៗទៅ សារធាតុផ្សេងៗ រួមទាំងសារធាតុតានីន ឬសារធាតុដែលស្រដៀងនឹងខ្លាញ់ កកកុញនៅក្នុងភ្នាសកោសិការបស់រុក្ខជាតិ ដែលនាំទៅដល់ការលេចធ្លាយ ឬការកកិតនៃជញ្ជាំងកោសិកា ការផ្លាស់ទីលំនៅនៃទឹក និងការស្លាប់នៃមាតិកាកោសិកា។ នៅចន្លោះជញ្ជាំងកោសិកានៃកោសិការុក្ខជាតិជិតខាងមានបន្ទះដូចចាហួយ - បន្ទះកណ្តាលដែលភ្ជាប់ពួកវាជាមួយគ្នា និងស៊ីម៉ង់ដល់រាងកាយរុក្ខជាតិទាំងមូល។ ពួកវាត្រូវបានបំផ្លាញតែនៅក្នុងដំណើរការនៃការទុំផ្លែឈើ និងនៅពេលដែលស្លឹកធ្លាក់ចុះ។

ជញ្ជាំងកោសិកានៃកោសិកាផ្សិតត្រូវបានបង្កើតឡើង ឈីទីន- កាបូអ៊ីដ្រាតដែលមានអាសូត។ ពួកវារឹងមាំគ្រប់គ្រាន់ និងជាគ្រោងឆ្អឹងខាងក្រៅនៃកោសិកា ប៉ុន្តែនៅតែដូចជានៅក្នុងរុក្ខជាតិ ពួកគេការពារជំងឺ phagocytosis ។
នៅក្នុងបាក់តេរី ជញ្ជាំងកោសិកាផ្ទុកនូវកាបូអ៊ីដ្រាតជាមួយនឹងបំណែកនៃ peptides - mureinទោះជាយ៉ាងណាក៏ដោយ ខ្លឹមសាររបស់វាប្រែប្រួលយ៉ាងខ្លាំងនៅក្នុងក្រុមផ្សេងៗនៃបាក់តេរី។ នៅលើជញ្ជាំងកោសិកា សារធាតុ polysaccharides ផ្សេងទៀតក៏អាចត្រូវបានបញ្ចេញផងដែរ ដែលបង្កើតជាកន្សោម mucous ដែលការពារបាក់តេរីពីឥទ្ធិពលខាងក្រៅ។
សែលកំណត់រូបរាងរបស់កោសិកា បម្រើជាជំនួយមេកានិក អនុវត្តមុខងារការពារ ផ្តល់នូវលក្ខណៈសម្បត្តិ osmotic នៃកោសិកា កំណត់ការលាតសន្ធឹងនៃមាតិការស់នៅ និងការពារការដាច់នៃកោសិកា ដែលកើនឡើងដោយសារតែការហូរចូលនៃកោសិកា។ ទឹក។ លើសពីនេះ ទឹក និងសារធាតុដែលរលាយក្នុងវាយកឈ្នះលើជញ្ជាំងកោសិកាមុននឹងចូលទៅក្នុងស៊ីតូប្លាសស៊ីម ឬផ្ទុយទៅវិញនៅពេលទុកវាចោល ខណៈពេលដែលទឹកត្រូវបានដឹកជញ្ជូនតាមជញ្ជាំងកោសិកាលឿនជាងតាមរយៈស៊ីតូប្លាស។
ស៊ីតូប្លាស្មា
ស៊ីតូប្លាស្មាគឺជាផ្នែកខាងក្នុងនៃកោសិកា។ សរីរាង្គទាំងអស់នៃកោសិកា ស្នូល និងផលិតផលកាកសំណល់ផ្សេងៗត្រូវបានជ្រមុជនៅក្នុងវា។
cytoplasm ភ្ជាប់ផ្នែកទាំងអស់នៃកោសិកាជាមួយគ្នា ប្រតិកម្មមេតាប៉ូលីសជាច្រើនកើតឡើងនៅក្នុងវា។ cytoplasm ត្រូវបានបំបែកចេញពីបរិស្ថាន ហើយបែងចែកជាផ្នែកៗដោយភ្នាស ពោលគឺកោសិកាមានរចនាសម្ព័ន្ធភ្នាស។ វាអាចស្ថិតនៅក្នុងរដ្ឋពីរ - សូលនិងជែល។ សូល។- នេះគឺជាស្ថានភាពពាក់កណ្តាលរាវ ស្រដៀងនឹងចាហួយនៃ cytoplasm ដែលដំណើរការសំខាន់ៗដំណើរការយ៉ាងសកម្មបំផុត និង ជែល- ស្ថានភាព gelatinous ក្រាស់ ដែលរារាំងលំហូរនៃប្រតិកម្មគីមី និងការដឹកជញ្ជូនសារធាតុ។
ផ្នែករាវនៃ cytoplasm ដោយគ្មានសរីរាង្គត្រូវបានគេហៅថា hyaloplasm. Hyaloplasm ឬ cytosol គឺជាសូលុយស្យុង colloidal ដែលក្នុងនោះមានប្រភេទនៃការព្យួរនៃភាគល្អិតធំល្មម ដូចជាប្រូតេអ៊ីន ដែលព័ទ្ធជុំវិញដោយ dipoles នៃម៉ូលេគុលទឹក។ ភាពច្របូកច្របល់នៃការព្យួរនេះមិនកើតឡើងដោយសារតែការពិតដែលថាពួកគេមានបន្ទុកដូចគ្នានិង repel គ្នាទៅវិញទៅមក។
សរីរាង្គ
សរីរាង្គ- ទាំងនេះគឺជាសមាសធាតុអចិន្ត្រៃយ៍នៃក្រឡាដែលបំពេញមុខងារជាក់លាក់។
អាស្រ័យលើលក្ខណៈរចនាសម្ព័ន្ធពួកវាត្រូវបានបែងចែកទៅជាភ្នាសនិងមិនមែនភ្នាស។ ភ្នាសនៅក្នុងវេន organelles ត្រូវបានគេហៅថា single-membrane (endoplasmic reticulum, Golgi complex and lysosomes) ឬ two-membrane (mitochondria, plastids and nucleus)។ មិនមែនភ្នាសសរីរាង្គគឺជា ribosomes, microtubules, microfilaments និងមជ្ឈមណ្ឌលកោសិកា។ ក្នុងចំណោមសរីរាង្គដែលបានរាយបញ្ជី មានតែ ribosomes ប៉ុណ្ណោះដែលមាននៅក្នុង prokaryotes ។
រចនាសម្ព័ន្ធនិងមុខងារនៃស្នូល។ ស្នូល- សរីរាង្គដែលមានភ្នាសពីរធំស្ថិតនៅកណ្តាលកោសិកា ឬនៅបរិវេណរបស់វា។ ទំហំនៃស្នូលអាចប្រែប្រួលក្នុងរង្វង់ 3-35 មីក្រូ។ រូបរាងរបស់ស្នូលច្រើនតែមានរាងស្វ៊ែរ ឬរាងអេលីប ប៉ុន្តែក៏មានស្នូលរាងជាកំណាត់ រាងពងក្រពើ រាងសណ្តែក lobed និងសូម្បីតែស្នូលដែលបែងចែក។ អ្នកស្រាវជ្រាវខ្លះជឿថារូបរាងនៃស្នូលត្រូវគ្នាទៅនឹងរូបរាងនៃកោសិកាខ្លួនឯង។
កោសិកាភាគច្រើនមានស្នូលតែមួយ ប៉ុន្តែឧទាហរណ៍ កោសិកាថ្លើម និងបេះដូងអាចមានពីរ ហើយក្នុងចំនួនណឺរ៉ូនរហូតដល់ 15។ សរសៃសាច់ដុំគ្រោងឆ្អឹងជាធម្មតាមានស្នូលជាច្រើន ប៉ុន្តែពួកវាមិនមែនជាកោសិកាក្នុងន័យពេញលេញនៃ ពាក្យ, ចាប់តាំងពីពួកគេត្រូវបានបង្កើតឡើងនៅក្នុងលទ្ធផលនៃការលាយបញ្ចូលគ្នានៃកោសិកាជាច្រើន។
ស្នូលត្រូវបានព័ទ្ធជុំវិញ ស្រោមសំបុត្រនុយក្លេអ៊ែរហើយទំហំខាងក្នុងរបស់វាត្រូវបានបំពេញ ទឹកនុយក្លេអ៊ែរ, ឬ nucleoplasm (karyoplasm)នៅក្នុងការដែលត្រូវបានជ្រមុជ ក្រូម៉ាទីននិង ស្នូល. ស្នូលដំណើរការមុខងារសំខាន់ៗដូចជាការផ្ទុក និងការបញ្ជូនព័ត៌មានតំណពូជ ក៏ដូចជាការគ្រប់គ្រងសកម្មភាពសំខាន់ៗរបស់កោសិកា។

តួនាទីរបស់ស្នូលក្នុងការបញ្ជូនព័ត៌មានតំណពូជត្រូវបានបង្ហាញយ៉ាងជឿជាក់ក្នុងការពិសោធន៍ជាមួយ acetabularia សារាយបៃតង។ នៅក្នុងកោសិកាយក្សតែមួយដែលឈានដល់ប្រវែង 5 សង់ទីម៉ែត្រ មួក ជើង និង rhizoid ត្រូវបានសម្គាល់។ ជាងនេះទៅទៀត វាមានស្នូលតែមួយដែលមានទីតាំងនៅក្នុង rhizoid ។ នៅទសវត្សរ៍ឆ្នាំ 1930 I. Hemmerling បានប្តូរស្នូលនៃប្រភេទសត្វ acetabularia មួយប្រភេទដែលមានពណ៌បៃតងទៅជា rhizoid នៃប្រភេទផ្សេងទៀតដែលមានពណ៌ត្នោត ដែលក្នុងនោះស្នូលត្រូវបានដកចេញ។ មួយសន្ទុះក្រោយមក រុក្ខជាតិដែលមានស្នូលស្ទូងបានដុះពន្លកថ្មី ដូចជាអ្នកផ្តល់ជំនួយនៃស្នូលសារាយ។ ក្នុងពេលជាមួយគ្នានោះ ដើម ឬដើមដែលបំបែកចេញពី rhizoid ដែលមិនមានស្នូល បានស្លាប់បន្ទាប់ពីពេលខ្លះ។

ស្រោមសំបុត្រនុយក្លេអ៊ែរវាត្រូវបានបង្កើតឡើងដោយភ្នាសពីរ - ខាងក្រៅនិងខាងក្នុងរវាងវាមានចន្លោះមួយ។ ចន្លោះ interemembrane ទាក់ទងជាមួយបែហោងធ្មែញនៃ reticulum endoplasmic រដុប ហើយភ្នាសខាងក្រៅនៃស្នូលអាចផ្ទុក ribosomes ។ ស្រោមសំបុត្រនុយក្លេអ៊ែរត្រូវបានជ្រាបចូលទៅក្នុងរន្ធញើសជាច្រើន គែមដោយប្រូតេអ៊ីនពិសេស។ សារធាតុត្រូវបានដឹកជញ្ជូនតាមរន្ធញើស៖ ប្រូតេអ៊ីនចាំបាច់ (រួមទាំងអង់ស៊ីម) អ៊ីយ៉ុង នុយក្លេអូទីត និងសារធាតុផ្សេងទៀតចូលទៅក្នុងស្នូល ហើយម៉ូលេគុល RNA ប្រូតេអ៊ីនកាកសំណល់ អនុផ្នែកនៃ ribosomes ទុកវាចោល។ ដូច្នេះមុខងារនៃស្រោមសំបុត្រនុយក្លេអ៊ែរគឺជាការបំបែកមាតិកានៃស្នូលចេញពី cytoplasm ក៏ដូចជាបទប្បញ្ញត្តិនៃការរំលាយអាហាររវាងស្នូលនិង cytoplasm ។
នុយក្លេអូផ្លាមហៅថាមាតិកានៃស្នូល ដែលក្រូម៉ាទីន និងនុយក្លេអូលត្រូវបានជ្រមុជ។ វាជាសូលុយស្យុង colloidal ដែលមានសារៈធាតុគីមីនៃ cytoplasm ។ អង់ស៊ីមនៃ nucleoplasm ជំរុញការផ្លាស់ប្តូរអាស៊ីតអាមីណូ នុយក្លេអូទីត ប្រូតេអ៊ីន។ល។ nucleoplasm ត្រូវបានភ្ជាប់ទៅនឹង hyaloplasm តាមរយៈរន្ធញើសនុយក្លេអ៊ែរ។ មុខងារនៃ nucleoplasm ដូចជា hyaloplasm គឺដើម្បីធានាការភ្ជាប់គ្នានៃសមាសធាតុរចនាសម្ព័ន្ធទាំងអស់នៃ nucleus និងការអនុវត្តប្រតិកម្មអង់ស៊ីមមួយចំនួន។
ក្រូម៉ាទីនហៅថា បណ្តុំនៃសរសៃស្តើង និងគ្រាប់ដែលដាក់នៅក្នុង nucleoplasm ។ វាអាចត្រូវបានគេរកឃើញដោយការប្រឡាក់ដោយសារសន្ទស្សន៍ចំណាំងផ្លាតនៃ chromatin និង nucleoplasm គឺប្រហែលដូចគ្នា។ សមាសធាតុ filamentous នៃ chromatin ត្រូវបានគេហៅថា euchromatin, និង granular ថ្នាំ heterochromatin. Euchromatin ត្រូវបានបង្រួមយ៉ាងទន់ខ្សោយ ចាប់តាំងពីព័ត៌មានតំណពូជត្រូវបានអានពីវា ខណៈពេលដែល heterochromatin spiralized កាន់តែច្រើនគឺអសកម្មហ្សែន។
Chromatin គឺជាការកែប្រែរចនាសម្ព័ន្ធនៃក្រូម៉ូសូមនៅក្នុងស្នូលដែលមិនបែងចែក។ ដូច្នេះ ក្រូម៉ូសូមមានវត្តមានជានិច្ចនៅក្នុងស្នូល មានតែស្ថានភាពរបស់វាប្រែប្រួលអាស្រ័យលើមុខងារដែលស្នូលធ្វើនៅពេលនេះ។
Chromatin មានជាចម្បងនៃ nucleoproteins (deoxyribonucleoproteins និង ribonucleoproteins) ក៏ដូចជាអង់ស៊ីម ដែលសំខាន់បំផុតត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការសំយោគអាស៊ីត nucleic និងសារធាតុមួយចំនួនទៀត។
មុខងាររបស់ chromatin ទីមួយមាននៅក្នុងការសំយោគអាស៊ីត nucleic ជាក់លាក់សម្រាប់សារពាង្គកាយដែលបានផ្តល់ឱ្យ ដែលដឹកនាំការសំយោគប្រូតេអ៊ីនជាក់លាក់ និងទីពីរក្នុងការផ្ទេរលក្ខណៈសម្បត្តិតំណពូជពីកោសិកាម្តាយទៅកោសិកាកូនស្រី ដែលខ្សែស្រឡាយ chromatin គឺ ខ្ចប់ចូលទៅក្នុងក្រូម៉ូសូមក្នុងអំឡុងពេលបែងចែក។
ស្នូល- រាងស្វ៊ែរ អាចមើលឃើញយ៉ាងច្បាស់នៅក្រោមមីក្រូទស្សន៍ដែលមានអង្កត់ផ្ចិត 1-3 មីក្រូ។ វាត្រូវបានបង្កើតឡើងនៅក្នុងតំបន់ក្រូម៉ាទីនដែលអ៊ិនកូដព័ត៌មានអំពីរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន rRNA និង ribosome ។ នុយក្លេអូលនៅក្នុងស្នូលជារឿយៗគឺមួយ ប៉ុន្តែនៅក្នុងកោសិកាទាំងនោះដែលដំណើរការសំខាន់ដែលពឹងផ្អែកខ្លាំងកើតឡើង វាអាចមាននុយក្លេអូលីពីរ ឬច្រើន។ មុខងាររបស់ nucleoli គឺជាការសំយោគនៃ rRNA និងការប្រមូលផ្តុំនៃអនុក្រុម ribosome ដោយរួមបញ្ចូលគ្នានូវ rRNA ជាមួយនឹងប្រូតេអ៊ីនដែលចេញមកពី cytoplasm ។
មីតូខុនឌ្រី- សរីរៈសរីរាង្គពីរនៃរាងមូល រាងពងក្រពើ ឬរាងមូល ទោះបីរាងជារង្វង់ក៏ត្រូវបានរកឃើញដែរ (នៅក្នុងមេជីវិតឈ្មោល)។ Mitochondria មានអង្កត់ផ្ចិតរហូតដល់ 1µm និងមានប្រវែងរហូតដល់ 7µm។ ចន្លោះខាងក្នុង mitochondria ត្រូវបានបំពេញដោយម៉ាទ្រីស។ ម៉ាទ្រីសវាគឺជាសារធាតុសំខាន់នៃ mitochondria ។ ម៉ូលេគុល DNA រាងជារង្វង់ និង ribosomes ត្រូវបានជ្រមុជនៅក្នុងវា។ ភ្នាសខាងក្រៅនៃ mitochondria គឺរលោង និងមិនអាចជ្រាបចូលបានទៅនឹងសារធាតុជាច្រើន។ ភ្នាសខាងក្នុងមានការរីកចម្រើន - គ្រីស្តាដែលបង្កើនផ្ទៃនៃភ្នាសសម្រាប់ប្រតិកម្មគីមីកើតឡើង។ នៅលើផ្ទៃនៃភ្នាសមានស្រទាប់ប្រូតេអ៊ីនជាច្រើនដែលបង្កើតបានជាខ្សែសង្វាក់ផ្លូវដង្ហើម ក៏ដូចជាអង់ស៊ីមរាងផ្សិតនៃ ATP synthetase ។ នៅក្នុង mitochondria ដំណាក់កាលនៃការដកដង្ហើមតាមបែប aerobic កើតឡើងក្នុងអំឡុងពេលដែល ATP ត្រូវបានសំយោគ។

ផ្លាស្ទីត- សរីរាង្គភ្នាសពីរធំ លក្ខណៈសម្រាប់តែកោសិការុក្ខជាតិ។ ចន្លោះខាងក្នុងនៃផ្លាស្ទីតត្រូវបានបំពេញ ស្ត្រូម៉ា, ឬ ម៉ាទ្រីស. នៅក្នុង stroma មានប្រព័ន្ធដែលត្រូវបានអភិវឌ្ឍច្រើនឬតិចនៃភ្នាសភ្នាស - thylakoidsដែលត្រូវបានប្រមូលជាគំនរ - គ្រាប់ធញ្ញជាតិក៏ដូចជាម៉ូលេគុល DNA រាងជារង្វង់ និង ribosomes របស់វា។ ផ្លាស្ទីតមានបួនប្រភេទសំខាន់ៗ៖ ក្លរ៉ូផ្លាស្ទីត ក្រូម៉ូប្លាស លីកូប្លាស និងប្រូផ្លាស្ទីត។
Chloroplast- ទាំងនេះគឺជាផ្លាស្ទីតពណ៌បៃតងដែលមានអង្កត់ផ្ចិត 3-10 មីក្រូ ដែលអាចមើលឃើញយ៉ាងច្បាស់នៅក្រោមមីក្រូទស្សន៍។ ពួកវាត្រូវបានរកឃើញតែនៅក្នុងផ្នែកពណ៌បៃតងនៃរុក្ខជាតិ - ស្លឹក ដើមវ័យក្មេង ផ្កា និងផ្លែឈើ។ Chloroplast ភាគច្រើនមានរាងជារាងពងក្រពើ ឬរាងអេលីប ប៉ុន្តែក៏អាចមានរាងជាពែង រាងជាវង់ និងសូម្បីតែរាងពងក្រពើ។ ចំនួន chloroplasts ក្នុងកោសិកាមួយជាមធ្យមពី 10 ទៅ 100 បំណែក។ ទោះយ៉ាងណាក៏ដោយ ជាឧទាហរណ៍ នៅក្នុងសារាយខ្លះវាអាចជាមួយ មានទំហំគួរឱ្យកត់សម្គាល់ និងរូបរាងស្មុគស្មាញ - បន្ទាប់មកវាត្រូវបានគេហៅថា ក្រូម៉ាតូហ្វ័រ. ក្នុងករណីផ្សេងទៀតចំនួន chloroplasts អាចឈានដល់រាប់រយ ខណៈទំហំរបស់វាតូច។ ពណ៌នៃ chloroplasts គឺដោយសារតែសារធាតុពណ៌សំខាន់នៃការធ្វើរស្មីសំយោគ - ក្លរ៉ូហ្វីលទោះបីជាពួកគេមានសារធាតុពណ៌បន្ថែមក៏ដោយ - carotenoids. Carotenoids ក្លាយជាគួរឱ្យកត់សម្គាល់តែនៅក្នុងរដូវស្លឹកឈើជ្រុះនៅពេលដែល chlorophyll នៅក្នុងស្លឹកចាស់ត្រូវបានបំផ្លាញ។ មុខងារសំខាន់របស់ chloroplasts គឺរស្មីសំយោគ។ ប្រតិកម្មពន្លឺនៃរស្មីសំយោគកើតឡើងនៅលើភ្នាស thylakoid ដែលម៉ូលេគុល chlorophyll ត្រូវបានជួសជុល ហើយប្រតិកម្មងងឹតកើតឡើងនៅក្នុង stroma ដែលមានអង់ស៊ីមជាច្រើន។
Chromoplastsមានពណ៌លឿង ពណ៌ទឹកក្រូច និងក្រហមដែលមានសារធាតុពណ៌ carotenoid ។ រូបរាងរបស់ chromoplasts ក៏អាចប្រែប្រួលគួរឱ្យកត់សម្គាល់ផងដែរ៖ ពួកវាមានរាងជាបំពង់ ស្វ៊ែរ គ្រីស្តាល់។
លីកូប្លាស- ទាំងនេះគឺជាផ្លាស្ទីតពណ៌ស ឬគ្មានពណ៌ ដែលភាគច្រើនមានរាងមូល ឬរាងពងក្រពើ។ ពួកវាមានជាទូទៅនៅក្នុងផ្នែកដែលមិនមែនជាសារធាតុសំយោគនៃរុក្ខជាតិ ដូចជាសំបកស្លឹក មើមដំឡូងជាដើម ពួកវាផ្ទុកសារធាតុចិញ្ចឹម ដែលភាគច្រើនជាម្សៅ ប៉ុន្តែនៅក្នុងរុក្ខជាតិខ្លះវាអាចជាប្រូតេអ៊ីន ឬប្រេង។
Plastids ត្រូវបានបង្កើតឡើងនៅក្នុងកោសិការុក្ខជាតិពី proplastids ដែលមានវត្តមានរួចហើយនៅក្នុងកោសិកានៃជាលិកាអប់រំ និងជាសាកសពតូចៗដែលមានភ្នាសពីរ។ នៅដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍ ផ្លាស្ទីតផ្សេងៗគ្នាអាចប្រែជាគ្នាទៅវិញទៅមក៖ នៅពេលដែលប៉ះនឹងពន្លឺ ដុំសាច់នៃមើមដំឡូង និងក្រូម៉ូប្លាសនៃឫសការ៉ុតប្រែទៅជាពណ៌បៃតង។
Plastids និង mitochondria ត្រូវបានគេហៅថាសរីរាង្គកោសិកាពាក់កណ្តាលស្វយ័ត ដោយសារពួកវាមានម៉ូលេគុល DNA និង ribosomes ផ្ទាល់របស់ពួកគេ អនុវត្តការសំយោគប្រូតេអ៊ីន និងបែងចែកដោយឯករាជ្យនៃការបែងចែកកោសិកា។ លក្ខណៈពិសេសទាំងនេះត្រូវបានពន្យល់ដោយប្រភពដើមពីសារពាង្គកាយ prokaryotic unicellular ។ ទោះជាយ៉ាងណាក៏ដោយ "ឯករាជ្យ" នៃ mitochondria និង plastids ត្រូវបានកំណត់ ចាប់តាំងពី DNA របស់ពួកគេមានហ្សែនតិចតួចពេកសម្រាប់អត្ថិភាពដោយឥតគិតថ្លៃ ខណៈដែលព័ត៌មានដែលនៅសល់ត្រូវបានអ៊ិនកូដនៅក្នុងក្រូម៉ូសូមនៃស្នូលដែលអនុញ្ញាតឱ្យវាគ្រប់គ្រងសរីរាង្គទាំងនេះ។
រ៉េទីកូឡាម Endoplasmic (ER), ឬ រីទីគូល endoplasmic (ER)គឺជាសរីរាង្គដែលមានភ្នាសតែមួយ ដែលជាបណ្តាញនៃបែហោងធ្មែញភ្នាស និងបំពង់ដែលកាន់កាប់រហូតដល់ 30% នៃមាតិកានៃ cytoplasm ។ អង្កត់ផ្ចិតនៃបំពង់ ER គឺប្រហែល 25-30 nm ។ EPS មានពីរប្រភេទ - រដុប និងរលោង។ XPS រដុបផ្ទុក ribosomes និងជាកន្លែងដែលប្រូតេអ៊ីនត្រូវបានសំយោគ។ EPS រលោងដោយគ្មាន ribosomes ។ មុខងាររបស់វាគឺការសំយោគ lipids និងកាបូអ៊ីដ្រាត ក៏ដូចជាការដឹកជញ្ជូន ការផ្ទុក និងការចោលសារធាតុពុល។ វាត្រូវបានបង្កើតឡើងជាពិសេសនៅក្នុងកោសិកាទាំងនោះដែលដំណើរការមេតាបូលីសដែលពឹងផ្អែកខ្លាំងកើតឡើង ឧទាហរណ៍នៅក្នុងកោសិកាថ្លើម - hepatocytes - និងសរសៃសាច់ដុំគ្រោងឆ្អឹង។ សារធាតុដែលត្រូវបានសំយោគនៅក្នុង EPS ត្រូវបានបញ្ជូនទៅឧបករណ៍ Golgi ។ នៅក្នុង ER ភ្នាសកោសិកាក៏ត្រូវបានប្រមូលផ្តុំផងដែរ ប៉ុន្តែការបង្កើតរបស់វាត្រូវបានបញ្ចប់នៅក្នុងបរិធាន Golgi ។
ឧបករណ៍ហ្គោលហ្គី,ឬ ហ្គោលជីស្មុគ្រស្មាញគឺជាសរីរាង្គដែលមានភ្នាសតែមួយ បង្កើតឡើងដោយប្រព័ន្ធនៃអាងទឹក បំពង់ និង vesicles ចេញពីពួកវា។ ឯកតារចនាសម្ព័ន្ធនៃបរិធាន Golgi គឺ ឌីតូសូម- ជង់នៃរថក្រោះមួយទៅកាន់បង្គោលមួយ ដែលសារធាតុពី ER មក ហើយពីប៉ូលទល់មុខ ដោយបានឆ្លងកាត់ការបំប្លែងមួយចំនួន ពួកវាត្រូវខ្ចប់ទៅជាពពុះ ហើយបញ្ជូនទៅផ្នែកផ្សេងទៀតនៃកោសិកា។ អង្កត់ផ្ចិតនៃធុងគឺប្រហែល 2 មីក្រូនហើយពពុះតូចៗមានប្រហែល 20-30 មីក្រូ។ មុខងារសំខាន់នៃស្មុគស្មាញ Golgi គឺការសំយោគសារធាតុមួយចំនួន និងការកែប្រែ (ផ្លាស់ប្តូរ) ប្រូតេអ៊ីន ខ្លាញ់ និងកាបូអ៊ីដ្រាតចេញពី ER ការបង្កើតភ្នាសចុងក្រោយ ក៏ដូចជាការដឹកជញ្ជូនសារធាតុតាមរយៈកោសិកា ការបន្តនៃ រចនាសម្ព័ន្ធរបស់វានិងការបង្កើត lysosomes ។ ឧបករណ៍ Golgi បានទទួលឈ្មោះរបស់ខ្លួនជាកិត្តិយសដល់អ្នកវិទ្យាសាស្ត្រអ៊ីតាលី Camillo Golgi ដែលបានរកឃើញសរីរាង្គនេះជាលើកដំបូង (1898) ។

លីសូសូម- សរីរាង្គភ្នាសតែមួយតូចរហូតដល់ 1 មីក្រូនក្នុងអង្កត់ផ្ចិតដែលមានអង់ស៊ីមអ៊ីដ្រូលីកដែលពាក់ព័ន្ធនឹងការរំលាយអាហារក្នុងកោសិកា។ ភ្នាសនៃ lysosomes គឺមិនអាចជ្រាបចូលបានយ៉ាងលំបាកសម្រាប់អង់ស៊ីមទាំងនេះ ដូច្នេះការអនុវត្តមុខងាររបស់ពួកគេដោយ lysosomes គឺត្រឹមត្រូវ និងកំណត់គោលដៅ។ ដូច្នេះ ពួកវាចូលរួមយ៉ាងសកម្មក្នុងដំណើរការនៃ phagocytosis បង្កើតជាបំពង់រំលាយអាហារ ហើយក្នុងករណីមានការអត់ឃ្លាន ឬខូចខាតដល់ផ្នែកខ្លះនៃកោសិកា ពួកវារំលាយពួកវាដោយមិនប៉ះពាល់ដល់អ្នកដទៃឡើយ។ ថ្មីៗនេះតួនាទីរបស់ lysosomes ក្នុងដំណើរការស្លាប់កោសិកាត្រូវបានរកឃើញ។
វ៉ាកូអូល។- បែហោងធ្មែញនៅក្នុង cytoplasm នៃកោសិការុក្ខជាតិ និងសត្វ ចងដោយភ្នាស និងពោរពេញទៅដោយរាវ។ សារធាតុរំលាយ និងកន្ត្រាក់ស្បូនត្រូវបានរកឃើញនៅក្នុងកោសិកាប្រូតូហ្សូន។ អតីតចូលរួមក្នុងដំណើរការនៃ phagocytosis ព្រោះវាបំបែកសារធាតុចិញ្ចឹម។ ក្រោយមកទៀតធានានូវការថែរក្សាតុល្យភាពទឹក - អំបិលដោយសារតែ osmoregulation ។ នៅក្នុងសត្វពហុកោសិកា vacuoles រំលាយអាហារត្រូវបានរកឃើញជាចម្បង។
នៅក្នុងកោសិការុក្ខជាតិ vacuoles តែងតែមានវត្តមាន ពួកគេត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសពិសេស និងពោរពេញទៅដោយកោសិកា។ ភ្នាសជុំវិញ vacuole គឺស្រដៀងគ្នានៅក្នុងសមាសភាពគីមី រចនាសម្ព័ន្ធ និងមុខងារនៃភ្នាសប្លាស្មា។ កោសិកាតំណាងឱ្យដំណោះស្រាយ aqueous នៃសារធាតុ inorganic និងសរីរាង្គផ្សេងៗ រួមទាំងអំបិលរ៉ែ អាស៊ីតសរីរាង្គ កាបូអ៊ីដ្រាត ប្រូតេអ៊ីន glycosides អាល់កាឡូអ៊ីត។ល។ vacuole អាចកាន់កាប់រហូតដល់ 90% នៃបរិមាណកោសិកា និងរុញស្នូលទៅបរិវេណ។ ផ្នែកនៃកោសិកានេះអនុវត្តមុខងារស្តុកទុក បញ្ចេញចោល osmotic ការពារ lysosomal និងមុខងារផ្សេងៗទៀត ដោយសារវាប្រមូលផ្តុំសារធាតុចិញ្ចឹម និងផលិតផលកាកសំណល់ វាផ្តល់ការផ្គត់ផ្គង់ទឹក និងរក្សារូបរាង និងបរិមាណនៃកោសិកា ហើយក៏មានអង់ស៊ីមសម្រាប់បំបែកសារធាតុជាច្រើនផងដែរ។ សមាសធាតុកោសិកា។ លើសពីនេះទៀតសារធាតុសកម្មជីវសាស្រ្តនៃ vacuoles អាចការពារសត្វជាច្រើនពីការស៊ីរុក្ខជាតិទាំងនេះ។ នៅក្នុងរុក្ខជាតិមួយចំនួនដោយសារតែការហើមនៃ vacuoles ការលូតលាស់កោសិកាកើតឡើងដោយការលាតសន្ធឹង។
Vacuoles ក៏មានវត្តមាននៅក្នុងកោសិកានៃផ្សិត និងបាក់តេរីមួយចំនួនដែរ ប៉ុន្តែនៅក្នុងផ្សិតពួកវាអនុវត្តមុខងារ osmoregulation ប៉ុណ្ណោះ ខណៈពេលដែលនៅក្នុង cyanobacteria ពួកវារក្សាភាពរឹងមាំ និងចូលរួមក្នុងដំណើរការនៃការ assimilation អាសូតពីខ្យល់។
រីបូសូម- សរីរាង្គដែលមិនមែនជាភ្នាសតូចៗដែលមានអង្កត់ផ្ចិត 15-20 មីក្រូដែលមានផ្នែករងពីរ - ធំនិងតូច។ អនុផ្នែក Eukaryotic ribosome ត្រូវបានប្រមូលផ្តុំនៅក្នុង nucleolus ហើយបន្ទាប់មកដឹកជញ្ជូនទៅកាន់ cytoplasm ។ ribosomes នៃ prokaryotes, mitochondria, និង plastids គឺតូចជាង eukaryotes ។ អនុផ្នែក Ribosome រួមមាន rRNA និងប្រូតេអ៊ីន។

ចំនួន ribosomes នៅក្នុងកោសិកាមួយអាចឡើងដល់រាប់សិបលាន៖ នៅក្នុង cytoplasm, mitochondria និង plastids ពួកវាស្ថិតក្នុងស្ថានភាពសេរី ហើយនៅលើ ER រដុប ក្នុងស្ថានភាពចង។ ពួកគេចូលរួមក្នុងការសំយោគប្រូតេអ៊ីនជាពិសេសពួកគេអនុវត្តដំណើរការនៃការបកប្រែ - ជីវសំយោគនៃខ្សែសង្វាក់ polypeptide នៅលើម៉ូលេគុល mRNA ។ នៅលើ ribosomes ឥតគិតថ្លៃ ប្រូតេអ៊ីននៃ hyaloplasm, mitochondria, plastids និងប្រូតេអ៊ីនផ្ទាល់នៃ ribosomes ត្រូវបានសំយោគ ខណៈដែលនៅលើ ribosomes ដែលភ្ជាប់ទៅនឹង ER រដុប ប្រូតេអ៊ីនត្រូវបានបកប្រែសម្រាប់ការបញ្ចេញចេញពីកោសិកា ការប្រមូលផ្តុំភ្នាស ការបង្កើត lysosomes និង vacuoles ។
Ribosomes អាចស្ថិតនៅក្នុង hyaloplasm ដោយឯកឯង ឬប្រមូលផ្តុំជាក្រុមជាមួយនឹងការសំយោគដំណាលគ្នានៃសង្វាក់ polypeptide ជាច្រើននៅលើ mRNA មួយ។ ក្រុមនៃ ribosomes ទាំងនេះត្រូវបានគេហៅថា polyribosomes, ឬ ប៉ូលីសូម។

microtubules- ទាំងនេះគឺជាសរីរាង្គដែលមិនមែនជាភ្នាសប្រហោងរាងស៊ីឡាំង ដែលជ្រាបចូលទៅក្នុង cytoplasm ទាំងមូលនៃកោសិកា។ អង្កត់ផ្ចិតរបស់ពួកគេគឺប្រហែល 25 nm កម្រាស់ជញ្ជាំងគឺ 6-8 nm ។ ពួកវាត្រូវបានបង្កើតឡើងដោយម៉ូលេគុលប្រូតេអ៊ីនជាច្រើន។ ទូប៊ូលីនដែលដំបូងបង្កើតបាន 13 ខ្សែស្រដៀងនឹងអង្កាំ ហើយបន្ទាប់មកប្រមូលផ្តុំទៅជា microtubule ។ Microtubules បង្កើតជា cytoplasmic reticulum ដែលផ្តល់រូបរាង និងបរិមាណកោសិកា ភ្ជាប់ភ្នាសប្លាស្មាជាមួយផ្នែកផ្សេងទៀតនៃកោសិកា ផ្តល់ការដឹកជញ្ជូនសារធាតុតាមរយៈកោសិកា ចូលរួមក្នុងចលនានៃកោសិកា និងសមាសធាតុ intracellular ក៏ដូចជានៅក្នុងការបែងចែក។ នៃសម្ភារៈហ្សែន។ ពួកវាជាផ្នែកមួយនៃមជ្ឈមណ្ឌលកោសិកានិងសរីរាង្គនៃចលនា - flagella និង cilia ។
មីក្រូហ្វីលឬ មីក្រូហ្វីលក៏ជាសរីរាង្គដែលមិនមែនជាភ្នាសដែរ ទោះជាយ៉ាងណាក៏ដោយ ពួកវាមានរាងជាសរសៃ ហើយមិនត្រូវបានបង្កើតឡើងដោយ tubulin ទេប៉ុន្តែ សកម្មភាព. ពួកគេចូលរួមក្នុងដំណើរការនៃការដឹកជញ្ជូនភ្នាស ការទទួលស្គាល់អន្តរកោសិកា ការបែងចែកកោសិកា cytoplasm និងនៅក្នុងចលនារបស់វា។ នៅក្នុងកោសិកាសាច់ដុំ អន្តរកម្មនៃ microfilaments actin ជាមួយសរសៃ myosin ផ្តល់នូវការកន្ត្រាក់។
Microtubules និង microfilaments បង្កើតជាគ្រោងខាងក្នុងនៃកោសិកា គ្រោងឆ្អឹង. វាគឺជាបណ្តាញស្មុគ្រស្មាញនៃសរសៃដែលផ្តល់ការគាំទ្រមេកានិចសម្រាប់ភ្នាសប្លាស្មា កំណត់រូបរាងរបស់កោសិកា ទីតាំងនៃសរីរាង្គកោសិកា និងចលនារបស់ពួកគេកំឡុងពេលបែងចែកកោសិកា។

មជ្ឈមណ្ឌលកោសិកា- សរីរាង្គដែលមិនមែនជាភ្នាសដែលមានទីតាំងនៅកោសិកាសត្វនៅជិតស្នូល; វាអវត្តមាននៅក្នុងកោសិការុក្ខជាតិ។ ប្រវែងរបស់វាគឺប្រហែល 0.2-0.3 µm និងអង្កត់ផ្ចិត 0.1-0.15 µm ។ មជ្ឈមណ្ឌលកោសិកាមានពីរ កណ្តាលដេកនៅក្នុងយន្តហោះកាត់កែងគ្នា និង រង្វង់រស្មីពី microtubules ។ centriole នីមួយៗត្រូវបានបង្កើតឡើងដោយក្រុម microtubules ចំនួន 9 ដែលប្រមូលបានជាបី ពោលគឺ triplets ។ មជ្ឈមណ្ឌលកោសិកាចូលរួមក្នុងការប្រមូលផ្តុំនៃ microtubules ការបែងចែកសម្ភារៈតំណពូជនៃកោសិកាក៏ដូចជាការបង្កើត flagella និង cilia ។

សរីរាង្គនៃចលនា។ Flagellaនិង ស៊ីលីយ៉ាគឺជាការរីកធំនៃកោសិកាដែលគ្របដណ្ដប់ដោយប្លាស្មា។ សរីរាង្គទាំងនេះផ្អែកលើ microtubules ប្រាំបួនគូដែលមានទីតាំងនៅតាមបរិវេណនិង microtubules ឥតគិតថ្លៃពីរនៅកណ្តាល។ Microtubules ត្រូវបានភ្ជាប់គ្នាទៅវិញទៅមកដោយប្រូតេអ៊ីនផ្សេងៗដែលធានានូវគម្លាតសម្របសម្រួលរបស់ពួកគេពីអ័ក្ស - លំយោល។ ការប្រែប្រួលគឺអាស្រ័យលើថាមពល ពោលគឺថាមពលនៃចំណងម៉ាក្រូនៃ ATP ត្រូវបានចំណាយលើដំណើរការនេះ។ ការស្ដារឡើងវិញនូវ flagella ដែលបាត់បង់ និង cilia គឺជាមុខងារមួយ។ សាកសព basal, ឬ kintosomesដែលមានទីតាំងនៅមូលដ្ឋានរបស់ពួកគេ។
ប្រវែងនៃ cilia គឺប្រហែល 10-15 nm និងប្រវែងនៃ flagella គឺ 20-50 microns ។ ដោយសារតែចលនាតម្រង់ទិសយ៉ាងតឹងរ៉ឹងនៃ flagella និង cilia មិនត្រឹមតែចលនារបស់សត្វ unicellular, spermatozoa ជាដើមត្រូវបានអនុវត្តប៉ុណ្ណោះទេ ប៉ុន្តែវាក៏ត្រូវបានសម្អាតផ្លូវដង្ហើមផងដែរ ស៊ុតផ្លាស់ទីតាមបំពង់ fallopian ដោយសារតែផ្នែកទាំងអស់នេះរបស់មនុស្ស។ រាងកាយត្រូវបានតម្រង់ជួរជាមួយ epithelium ciliated ។

ការរួមបញ្ចូល
ការរួមបញ្ចូល- ទាំងនេះគឺជាសមាសធាតុមិនអចិន្ត្រៃយ៍នៃកោសិកាដែលត្រូវបានបង្កើតឡើងនិងបាត់នៅក្នុងដំណើរជីវិតរបស់វា។ ទាំងនេះរួមមានទាំងសារធាតុបម្រុង ឧទាហរណ៍ គ្រាប់ម្សៅ ឬប្រូតេអ៊ីននៅក្នុងកោសិការុក្ខជាតិ គ្រាប់ glycogen នៅក្នុងកោសិកាសត្វ និងផ្សិត volutin ក្នុងបាក់តេរី ការធ្លាក់ចុះជាតិខ្លាញ់ក្នុងប្រភេទកោសិកាទាំងអស់ និងផលិតផលកាកសំណល់ ជាពិសេសសំណល់អាហារដែលមិនបានរំលាយជាលទ្ធផល។ នៃ phagocytosis ។ បង្កើតអ្វីដែលគេហៅថាសាកសពសំណល់។
ទំនាក់ទំនងនៃរចនាសម្ព័ន្ធ និងមុខងារនៃផ្នែក និងសរីរាង្គនៃកោសិកា គឺជាមូលដ្ឋាននៃភាពសុចរិតរបស់វា
ផ្នែកនីមួយៗនៃកោសិកា ម្យ៉ាងវិញទៀតគឺជារចនាសម្ព័ន្ធដាច់ដោយឡែកដែលមានរចនាសម្ព័ន្ធ និងមុខងារជាក់លាក់ ហើយម្យ៉ាងវិញទៀត ធាតុផ្សំនៃប្រព័ន្ធស្មុគស្មាញជាងនេះហៅថាកោសិកា។ ភាគច្រើននៃព័ត៌មានតំណពូជនៃកោសិកា eukaryotic ត្រូវបានប្រមូលផ្តុំនៅក្នុងស្នូល ប៉ុន្តែស្នូលខ្លួនវាមិនអាចធានាបាននូវការអនុវត្តរបស់វាបានទេ ដោយសារវាទាមទារយ៉ាងហោចណាស់ cytoplasm ដែលដើរតួជាសារធាតុសំខាន់ និង ribosomes ដែលការសំយោគនេះកើតឡើង។ . ribosomes ភាគច្រើនមានទីតាំងនៅលើ granular endoplasmic reticulum ពីកន្លែងដែលប្រូតេអ៊ីនត្រូវបានដឹកជញ្ជូនជាញឹកញាប់បំផុតទៅកាន់ Golgi complex ហើយបន្ទាប់មក បន្ទាប់ពីការកែប្រែទៅផ្នែកទាំងនោះនៃកោសិកាដែលពួកគេមានបំណង ឬត្រូវបានបញ្ចេញចេញ។ ការវេចខ្ចប់ភ្នាសនៃប្រូតេអ៊ីន និងកាបូអ៊ីដ្រាតអាចត្រូវបានបញ្ចូលទៅក្នុងភ្នាសសរីរាង្គ និងភ្នាស cytoplasmic ដោយធានានូវការបង្កើតឡើងវិញជាបន្តបន្ទាប់។ Lysosomes និង vacuoles ដែលបំពេញមុខងារសំខាន់បំផុតក៏ត្រូវបានភ្ជាប់ពី Golgi complex ផងដែរ។ ជាឧទាហរណ៍ បើគ្មាន lysosomes ទេ កោសិកានឹងប្រែក្លាយទៅជាប្រភេទនៃសំណល់នៃម៉ូលេគុល និងរចនាសម្ព័ន្ធយ៉ាងឆាប់រហ័ស។
ដំណើរការទាំងអស់នេះត្រូវការថាមពលដែលផលិតដោយ mitochondria និងនៅក្នុងរុក្ខជាតិផងដែរដោយ chloroplasts ។ ហើយទោះបីជាសរីរាង្គទាំងនេះមានលក្ខណៈស្វយ័តក៏ដោយ ដោយសារពួកគេមានម៉ូលេគុល DNA ផ្ទាល់របស់ពួកគេ ប្រូតេអ៊ីនមួយចំនួននៅតែត្រូវបានអ៊ិនកូដដោយហ្សែននុយក្លេអ៊ែរ និងសំយោគនៅក្នុង cytoplasm ។
ដូច្នេះ ក្រឡាគឺជាការរួបរួមដែលមិនអាចបំបែកបាននៃសមាសធាតុផ្សំរបស់វា ដែលនីមួយៗដំណើរការមុខងារពិសេសរៀងៗខ្លួន។
ការរំលាយអាហារ និងការបំប្លែងថាមពល គឺជាលក្ខណៈសម្បត្តិរបស់សារពាង្គកាយមានជីវិត។ ការរំលាយអាហារថាមពល និងប្លាស្ទិក ទំនាក់ទំនងរបស់ពួកគេ។ ដំណាក់កាលនៃការរំលាយអាហារថាមពល។ fermentation និងការដកដង្ហើម។ រស្មីសំយោគ សារៈសំខាន់របស់វា តួនាទីលោហធាតុ។ ដំណាក់កាលនៃការសំយោគរស្មីសំយោគ។ ប្រតិកម្មពន្លឺនិងងងឹតនៃរស្មីសំយោគទំនាក់ទំនងរបស់ពួកគេ។ គីមីវិទ្យា។ តួនាទីរបស់បាក់តេរីគីមីវិទ្យានៅលើផែនដី
ការរំលាយអាហារនិងការបំប្លែងថាមពល - លក្ខណៈសម្បត្តិនៃសារពាង្គកាយមានជីវិត
កោសិកាអាចត្រូវបានគេប្រដូចទៅនឹងរោងចក្រគីមីខ្នាតតូច ដែលប្រតិកម្មគីមីរាប់រយពាន់កើតឡើង។
មេតាបូលីស- សំណុំនៃការបំប្លែងសារជាតិគីមី សំដៅដល់ការអភិរក្ស និងការបន្តពូជដោយខ្លួនឯងនៃប្រព័ន្ធជីវសាស្រ្ត។
វារួមបញ្ចូលទាំងការទទួលទានសារធាតុចូលទៅក្នុងរាងកាយក្នុងអំឡុងពេលអាហាររូបត្ថម្ភនិងការដកដង្ហើម, ការរំលាយអាហារ intracellular, ឬ ការរំលាយអាហារក៏ដូចជាការបែងចែកផលិតផលចុងក្រោយនៃការរំលាយអាហារ។
មេតាបូលីសត្រូវបានភ្ជាប់ដោយ inextricably ជាមួយនឹងដំណើរការនៃការបំលែងថាមពលមួយប្រភេទទៅជាថាមពលមួយផ្សេងទៀត។ ឧទាហរណ៍ នៅក្នុងដំណើរការនៃការធ្វើរស្មីសំយោគ ថាមពលពន្លឺត្រូវបានរក្សាទុកក្នុងទម្រង់ជាថាមពលនៃចំណងគីមីនៃម៉ូលេគុលសរីរាង្គស្មុគស្មាញ ហើយនៅក្នុងដំណើរការនៃការដកដង្ហើម វាត្រូវបានបញ្ចេញ និងចំណាយលើការសំយោគនៃម៉ូលេគុលថ្មី ការងារមេកានិច និង osmotic ។ ត្រូវបានរលាយក្នុងទម្រង់នៃកំដៅ។ល។
លំហូរនៃប្រតិកម្មគីមីនៅក្នុងសារពាង្គកាយមានជីវិតត្រូវបានធានាដោយកាតាលីករជីវសាស្រ្តនៃធម្មជាតិប្រូតេអ៊ីន - អង់ស៊ីម, ឬ អង់ស៊ីម. ដូចកាតាលីករដទៃទៀតដែរ អង់ស៊ីមបង្កើនល្បឿនលំហូរនៃប្រតិកម្មគីមីនៅក្នុងកោសិកាដោយរាប់សិប និងរាប់រយរាប់ពាន់ដង ហើយជួនកាលថែមទាំងធ្វើឱ្យពួកវាអាចធ្វើទៅបាន ប៉ុន្តែមិនផ្លាស់ប្តូរទាំងធម្មជាតិ ឬលក្ខណៈសម្បត្តិនៃផលិតផលចុងក្រោយ (ផលិតផល) នៃប្រតិកម្ម និង កុំផ្លាស់ប្តូរខ្លួនឯង។ អង់ស៊ីមអាចជាប្រូតេអ៊ីនសាមញ្ញ និងស្មុគស្មាញ ដែលបន្ថែមលើផ្នែកប្រូតេអ៊ីន ក៏រួមបញ្ចូលផ្នែកដែលមិនមែនជាប្រូតេអ៊ីនផងដែរ - cofactor (coenzyme). ឧទាហរណ៍នៃអង់ស៊ីមគឺ amylase salivary ដែលបំបែកសារធាតុ polysaccharides ក្នុងអំឡុងពេលទំពារយូរ និង pepsin ដែលធានាដល់ការរំលាយប្រូតេអ៊ីននៅក្នុងក្រពះ។
អង់ស៊ីមខុសគ្នាពីកាតាលីករដែលមិនមានប្រូតេអ៊ីននៅក្នុងភាពជាក់លាក់ខ្ពស់នៃសកម្មភាពរបស់ពួកគេ ការកើនឡើងយ៉ាងខ្លាំងនៃអត្រាប្រតិកម្មជាមួយនឹងជំនួយរបស់ពួកគេ ក៏ដូចជាសមត្ថភាពក្នុងការគ្រប់គ្រងសកម្មភាពដោយការផ្លាស់ប្តូរលក្ខខណ្ឌប្រតិកម្ម ឬអន្តរកម្មជាមួយសារធាតុផ្សេងៗ។ លើសពីនេះ លក្ខខណ្ឌដែលដំណើរការកាតាលីករអង់ស៊ីមមានភាពខុសគ្នាខ្លាំងពីកត្តាដែលមិនមានអង់ស៊ីមកើតឡើង៖ សីតុណ្ហភាព $37°C$ គឺល្អបំផុតសម្រាប់ដំណើរការនៃអង់ស៊ីមក្នុងរាងកាយមនុស្ស សម្ពាធគួរតែនៅជិតបរិយាកាស និង $pH$ នៃឧបករណ៍ផ្ទុកអាចស្ទាក់ស្ទើរយ៉ាងខ្លាំង។ ដូច្នេះសម្រាប់អាមីឡាស បរិស្ថានអាល់កាឡាំងគឺចាំបាច់ ហើយសម្រាប់ pepsin ដែលជាអាស៊ីតមួយ។
យន្តការនៃសកម្មភាពនៃអង់ស៊ីមគឺដើម្បីកាត់បន្ថយថាមពលសកម្មនៃសារធាតុ (ស្រទាប់ខាងក្រោម) ដែលចូលទៅក្នុងប្រតិកម្មដោយសារតែការបង្កើតស្មុគស្មាញនៃអង់ស៊ីមស្រទាប់ខាងក្រោម។

ការរំលាយអាហារថាមពល និងប្លាស្ទិក ទំនាក់ទំនងរបស់ពួកគេ។
ការរំលាយអាហារមានដំណើរការពីរដែលកើតឡើងក្នុងពេលដំណាលគ្នានៅក្នុងកោសិកា៖ ការផ្លាស់ប្តូរប្លាស្ទិក និងថាមពល។
ការរំលាយអាហារផ្លាស្ទិច (anabolism, assimilation)គឺជាសំណុំនៃប្រតិកម្មសំយោគដែលទៅជាមួយការចំណាយនៃថាមពល ATP ។ នៅក្នុងដំណើរការនៃការរំលាយអាហារប្លាស្ទិកសារធាតុសរីរាង្គដែលចាំបាច់សម្រាប់កោសិកាត្រូវបានសំយោគ។ ឧទាហរណ៍នៃប្រតិកម្មផ្លាស់ប្តូរផ្លាស្ទិចគឺ ការសំយោគរស្មីសំយោគ ជីវសំយោគប្រូតេអ៊ីន និងការចម្លង DNA (ទ្វេដងដោយខ្លួនឯង)។
ការរំលាយអាហារថាមពល (catabolism, dissimilation)គឺជាសំណុំនៃប្រតិកម្មដែលបំបែកសារធាតុស្មុគស្មាញទៅជាសារធាតុសាមញ្ញជាង។ ជាលទ្ធផលនៃការរំលាយអាហារថាមពលថាមពលត្រូវបានបញ្ចេញដែលត្រូវបានរក្សាទុកក្នុងទម្រង់ ATP ។ ដំណើរការដ៏សំខាន់បំផុតនៃការបំប្លែងថាមពលគឺការដកដង្ហើម និងការ fermentation ។
ការផ្លាស់ប្តូរផ្លាស្ទិច និងថាមពលត្រូវបានភ្ជាប់គ្នាដោយ inextricably ចាប់តាំងពីសារធាតុសរីរាង្គត្រូវបានសំយោគនៅក្នុងដំណើរការនៃការផ្លាស់ប្តូរផ្លាស្ទិច ហើយនេះតម្រូវឱ្យមានថាមពល ATP ហើយនៅក្នុងដំណើរការនៃការបំប្លែងថាមពល សារធាតុសរីរាង្គត្រូវបានបំបែក ហើយថាមពលត្រូវបានបញ្ចេញ ដែលបន្ទាប់មកនឹងត្រូវចំណាយលើដំណើរការសំយោគ។ .
សារពាង្គកាយទទួលបានថាមពលក្នុងដំណើរការអាហារូបត្ថម្ភ ហើយបញ្ចេញវា ហើយបំប្លែងវាទៅជាទម្រង់ដែលអាចចូលដំណើរការបាន ជាចម្បងនៅក្នុងដំណើរការនៃការដកដង្ហើម។ យោងទៅតាមវិធីនៃអាហាររូបត្ថម្ភគ្រប់សារពាង្គកាយទាំងអស់ត្រូវបានបែងចែកទៅជា autotrophs និង heterotrophs ។ អូតូត្រូហ្វអាចសំយោគសារធាតុសរីរាង្គដោយឯករាជ្យពីអសរីរាង្គ និង heterotrophsប្រើសារធាតុសរីរាង្គដែលត្រៀមរួចជាស្រេច។
ដំណាក់កាលនៃការរំលាយអាហារថាមពល
ទោះបីជាមានភាពស្មុគ្រស្មាញនៃប្រតិកម្មមេតាប៉ូលីសថាមពលក៏ដោយ វាត្រូវបានបែងចែកតាមលក្ខខណ្ឌជាបីដំណាក់កាល៖ ត្រៀមលក្ខណៈ anaerobic (គ្មានអុកស៊ីសែន) និង aerobic (អុកស៊ីសែន)។
នៅលើ ដំណាក់កាលត្រៀមម៉ូលេគុលនៃ polysaccharides, lipids, ប្រូតេអ៊ីន, អាស៊ីត nucleic បំបែកទៅជារបស់សាមញ្ញជាង ឧទាហរណ៍ គ្លុយកូស glycerol និងអាស៊ីតខ្លាញ់ អាស៊ីតអាមីណូ nucleotides ជាដើម ដំណាក់កាលនេះអាចកើតឡើងដោយផ្ទាល់នៅក្នុងកោសិកា ឬក្នុងពោះវៀន ពីកន្លែងដែល សារធាតុបំបែកត្រូវបានចែកចាយជាមួយនឹងលំហូរឈាម។
ដំណាក់កាល anaerobicការបំប្លែងសារជាតិថាមពលត្រូវបានអមដោយការបំបែកបន្ថែមទៀតនៃសារធាតុ monomers នៃសមាសធាតុសរីរាង្គទៅកាន់ផលិតផលកម្រិតមធ្យមដ៏សាមញ្ញជាង ឧទាហរណ៍ អាស៊ីត pyruvic ឬ pyruvate ។ វាមិនតម្រូវឱ្យមានវត្តមានអុកស៊ីហ៊្សែនទេ ហើយសម្រាប់សារពាង្គកាយជាច្រើនដែលរស់នៅក្នុងដីល្បាប់នៃវាលភក់ ឬនៅក្នុងពោះវៀនរបស់មនុស្ស វាគឺជាមធ្យោបាយតែមួយគត់ដើម្បីទទួលបានថាមពល។ ដំណាក់កាល anaerobic នៃការរំលាយអាហារថាមពលកើតឡើងនៅក្នុង cytoplasm ។
សារធាតុផ្សេងៗអាចឆ្លងកាត់ការបំបែកដោយគ្មានអុកស៊ីហ្សែន ប៉ុន្តែគ្លុយកូសច្រើនតែជាស្រទាប់ខាងក្រោមនៃប្រតិកម្ម។ ដំណើរការនៃការបំបែកដោយគ្មានអុកស៊ីហ្សែនរបស់វាត្រូវបានគេហៅថា glycolysis. ក្នុងអំឡុងពេល glycolysis ម៉ូលេគុលគ្លុយកូសបាត់បង់អាតូមអ៊ីដ្រូសែនចំនួនបួន ពោលគឺវាត្រូវបានកត់សុី ហើយម៉ូលេគុលអាស៊ីត pyruvic ពីរម៉ូលេគុល ATP ពីរ និងម៉ូលេគុលពីរនៃនាវាផ្ទុកអ៊ីដ្រូសែនដែលកាត់បន្ថយ $NADH + H^(+)$ ត្រូវបានបង្កើតឡើង៖
$C_6H_(12)O_6 + 2H_3PO_4 + 2ADP + 2NAD → 2C_3H_4O_3 + 2ATP + 2NADH + H^(+) + 2H_2O$។
ការបង្កើត ATP ពី ADP កើតឡើងដោយសារតែការផ្ទេរដោយផ្ទាល់នៃ phosphate anion ពីជាតិស្ករ phosphorylated ពីមុន ហើយត្រូវបានគេហៅថា phosphorylation ស្រទាប់ខាងក្រោម។
ដំណាក់កាល Aerobicការផ្លាស់ប្តូរថាមពលអាចកើតឡើងបានតែនៅក្នុងវត្តមាននៃអុកស៊ីហ៊្សែន ខណៈពេលដែលសមាសធាតុកម្រិតមធ្យមដែលបង្កើតឡើងក្នុងដំណើរការនៃការបំបែកដោយគ្មានអុកស៊ីហ្សែនត្រូវបានកត់សុីទៅជាផលិតផលចុងក្រោយ (កាបូនឌីអុកស៊ីត និងទឹក) ហើយថាមពលភាគច្រើនដែលរក្សាទុកក្នុងចំណងគីមីនៃសមាសធាតុសរីរាង្គត្រូវបានបញ្ចេញ។ . វាចូលទៅក្នុងថាមពលនៃចំណងម៉ាក្រូនៃម៉ូលេគុល ATP ចំនួន 36 ។ ដំណាក់កាលនេះត្រូវបានគេហៅផងដែរ។ ការដកដង្ហើមជាលិកា. អវត្ដមាននៃអុកស៊ីហ៊្សែន សមាសធាតុកម្រិតមធ្យមត្រូវបានបំប្លែងទៅជាសារធាតុសរីរាង្គផ្សេងទៀត ដំណើរការហៅថា fermentation ។
ដង្ហើម
យន្តការនៃការដកដង្ហើមកោសិកាត្រូវបានបង្ហាញជាគ្រោងការណ៍នៅក្នុងរូបភព។

ការដកដង្ហើមតាមបែប Aerobic កើតឡើងនៅក្នុង mitochondria ខណៈពេលដែលអាស៊ីត pyruvic ដំបូងបាត់បង់អាតូមកាបូនមួយ ដែលត្រូវបានអមដោយការសំយោគនៃការកាត់បន្ថយមួយស្មើនឹង $NADH + H^(+)$ និងម៉ូលេគុលអាសេទីល កូអង់ស៊ីម A (អាសេទីល-CoA)៖
$C_3H_4O_3 + NAD + H~CoA → CH_3CO~CoA + NADH + H^(+) + CO_2$។
Acetyl-CoA នៅក្នុងម៉ាទ្រីស mitochondrial ត្រូវបានចូលរួមនៅក្នុងខ្សែសង្វាក់នៃប្រតិកម្មគីមី ដែលចំនួនសរុបត្រូវបានគេហៅថា វដ្ត Krebs (វដ្តអាស៊ីត tricarboxylic វដ្តអាស៊ីតនៃក្រូចឆ្មា) ក្នុងអំឡុងពេលនៃការផ្លាស់ប្តូរទាំងនេះ ម៉ូលេគុល ATP ពីរត្រូវបានបង្កើតឡើង acetyl-CoA ត្រូវបានកត់សុីទាំងស្រុងទៅជាកាបូនឌីអុកស៊ីត ហើយអ៊ីយ៉ុងអ៊ីដ្រូសែន និងអេឡិចត្រុងរបស់វាត្រូវបានភ្ជាប់ទៅនឹងក្រុមហ៊ុនដឹកជញ្ជូនអ៊ីដ្រូសែន $NADH + H^(+)$ និង $FADH_2$ ។ ក្រុមហ៊ុនដឹកជញ្ជូនដឹកជញ្ជូនប្រូតុងអ៊ីដ្រូសែន និងអេឡិចត្រុងទៅកាន់ភ្នាសខាងក្នុងនៃ mitochondria ដែលបង្កើតបានជាគ្រីស្តាល់។ ដោយមានជំនួយពីប្រូតេអ៊ីនក្រុមហ៊ុនដឹកជញ្ជូន ប្រូតុងអ៊ីដ្រូសែនត្រូវបានចាក់ចូលទៅក្នុងលំហរនៃភ្នាសខាងក្នុង ហើយអេឡិចត្រុងត្រូវបានផ្ទេរតាមខ្សែសង្វាក់ផ្លូវដង្ហើមនៃអង់ស៊ីមដែលមានទីតាំងនៅភ្នាសខាងក្នុងនៃ mitochondria ហើយត្រូវបានបោះចោលទៅលើអាតូមអុកស៊ីហ្សែន៖
$O_2+2e^(-) →O_2^-$ ។
វាគួរតែត្រូវបានគេកត់សម្គាល់ថាប្រូតេអ៊ីនមួយចំនួននៃខ្សែសង្វាក់ផ្លូវដង្ហើមមានជាតិដែកនិងស្ពាន់ធ័រ។
ពីចន្លោះ interemembrane ប្រូតុងអ៊ីដ្រូសែនត្រូវបានបញ្ជូនត្រឡប់ទៅម៉ាទ្រីស mitochondrial ដោយមានជំនួយពីអង់ស៊ីមពិសេស - ការសំយោគ ATP ហើយថាមពលដែលបានបញ្ចេញក្នុងករណីនេះត្រូវបានចំណាយលើការសំយោគម៉ូលេគុល ATP ចំនួន 34 ពីម៉ូលេគុលគ្លុយកូសនីមួយៗ។ ដំណើរការនេះត្រូវបានគេហៅថា ផូស្វ័រអុកស៊ីតកម្ម. នៅក្នុងម៉ាទ្រីស mitochondrial ប្រូតុងអ៊ីដ្រូសែនមានប្រតិកម្មជាមួយនឹងរ៉ាឌីកាល់អុកស៊ីសែនដើម្បីបង្កើតជាទឹក៖
$4H^(+)+O_2^- →2H_2O$ ។
សំណុំនៃប្រតិកម្មនៃការដកដង្ហើមអុកស៊ីសែនអាចត្រូវបានបង្ហាញដូចខាងក្រោម:
$2C_3H_4O_3 + 6O_2 + 36H_3PO_4 + 36ADP → 6CO_2 + 38H_2O + 36ATP.$
សមីការដង្ហើមទូទៅមើលទៅដូចនេះ៖
$C_6H_(12)O_6 + 6O_2 + 38H_3PO_4 + 38ADP → 6CO_2 + 40H_2O + 38ATP.$
ការ fermentation
អវត្ដមាននៃអុកស៊ីសែនឬកង្វះរបស់វា fermentation កើតឡើង។ ការ fermentation គឺជាវិធីវិវត្តន៍មុនគេក្នុងការទទួលបានថាមពលជាងការដកដង្ហើម ប៉ុន្តែវាមានផលចំណេញតិចជាងថាមពល ដោយសារតែការ fermentation ផលិតសារធាតុសរីរាង្គដែលនៅតែសម្បូរថាមពល។ មានប្រភេទសំខាន់ៗជាច្រើននៃការ fermentation៖ អាស៊ីតឡាក់ទិក ជាតិអាល់កុល អាស៊ីតអាសេទិក ជាដើម។ ដូច្នេះនៅក្នុងសាច់ដុំគ្រោងឆ្អឹង ក្នុងករណីដែលគ្មានអុកស៊ីហ្សែនក្នុងអំឡុងពេល fermentation អាស៊ីត pyruvic ត្រូវបានកាត់បន្ថយទៅជាអាស៊ីតឡាក់ទិក ខណៈដែលសមមូលកាត់បន្ថយដែលបានបង្កើតពីមុនត្រូវបានប្រើប្រាស់ និង នៅសល់តែម៉ូលេគុល ATP ពីរប៉ុណ្ណោះ៖
$2C_3H_4O_3 + 2NADH + H^(+) → 2C_3H_6O_3 + 2NAD$។
ក្នុងអំឡុងពេលនៃការ fermentation ដោយមានជំនួយពីផ្សិតផ្សិត អាស៊ីត pyruvic នៅក្នុងវត្តមាននៃអុកស៊ីសែនប្រែទៅជាអេទីលអាល់កុលនិងកាបូនម៉ូណូអុកស៊ីត (IV)៖
$C_3H_4O_3 + NADH + H^(+) → C_2H_5OH + CO_2 + NAD^(+)$ ។
ក្នុងអំឡុងពេល fermentation ដោយមានជំនួយពី microorganisms អាស៊ីត pyruvic ក៏អាចបង្កើត acetic, butyric, formic acids ជាដើម។
ATP ដែលទទួលបានជាលទ្ធផលនៃការបំប្លែងថាមពលត្រូវបានប្រើប្រាស់នៅក្នុងកោសិកាសម្រាប់ប្រភេទផ្សេងៗនៃការងារ៖ គីមី, osmotic, អគ្គិសនី, មេកានិច និងបទប្បញ្ញត្តិ។ ការងារគីមីមាននៅក្នុងការសំយោគជីវសាស្រ្តនៃប្រូតេអ៊ីន ខ្លាញ់ កាបូអ៊ីដ្រាត អាស៊ីត nucleic និងសមាសធាតុសំខាន់ៗផ្សេងទៀត។ ការងារ Osmotic រួមមានដំណើរការនៃការស្រូបយកដោយកោសិកា និងការដកយកចេញពីវានៃសារធាតុដែលមាននៅក្នុងចន្លោះ extracellular ក្នុងកំហាប់ធំជាងនៅក្នុងកោសិកា។ ការងារអគ្គិសនីគឺទាក់ទងយ៉ាងជិតស្និទ្ធទៅនឹងការងារ osmotic ព្រោះវាជាលទ្ធផលនៃចលនានៃភាគល្អិតដែលមានបន្ទុកតាមរយៈភ្នាសដែលបន្ទុកនៃភ្នាសត្រូវបានបង្កើតឡើង ហើយលក្ខណៈសម្បត្តិនៃការរំភើប និងចរន្តត្រូវបានទទួល។ ការងារមេកានិកត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងចលនានៃសារធាតុ និងរចនាសម្ព័ន្ធនៅខាងក្នុងកោសិកា ក៏ដូចជាកោសិកាទាំងមូល។ ការងារនិយតកម្មរួមមានដំណើរការទាំងអស់ដែលមានបំណងសម្របសម្រួលដំណើរការនៅក្នុងក្រឡា។
រស្មីសំយោគ សារៈសំខាន់របស់វា តួនាទីលោហធាតុ
រស្មីសំយោគហៅថាដំណើរការនៃការបំលែងថាមពលពន្លឺទៅជាថាមពលនៃចំណងគីមីនៃសមាសធាតុសរីរាង្គដោយមានការចូលរួមពីក្លរ៉ូហ្វីល។
ជាលទ្ធផលនៃការធ្វើរស្មីសំយោគ សារធាតុសរីរាង្គប្រហែល 150 ពាន់លានតោន និងប្រហែល 200 ពាន់លានតោននៃអុកស៊ីសែនត្រូវបានផលិតជារៀងរាល់ឆ្នាំ។ ដំណើរការនេះធានាដល់ចរាចរកាបូននៅក្នុងជីវមណ្ឌល ការពារការប្រមូលផ្តុំកាបូនឌីអុកស៊ីត និងដោយហេតុនេះការពារការកើតឡើងនៃឥទ្ធិពលផ្ទះកញ្ចក់ និងការឡើងកំដៅផែនដី។ សារធាតុសរីរាង្គដែលបានបង្កើតឡើងជាលទ្ធផលនៃការធ្វើរស្មីសំយោគមិនត្រូវបានប្រើប្រាស់ទាំងស្រុងដោយសារពាង្គកាយផ្សេងទៀតទេ ដែលជាផ្នែកសំខាន់នៃពួកវាបង្កើតបានជាប្រាក់បញ្ញើរ៉ែ (ធ្យូងថ្មរឹង និងពណ៌ត្នោត ប្រេង) ក្នុងរយៈពេលរាប់លានឆ្នាំ។ ថ្មីៗនេះ ប្រេង rapeseed (" biodiesel") និងអាល់កុលដែលទទួលបានពីសំណល់រុក្ខជាតិក៏ត្រូវបានគេប្រើជាឥន្ធនៈផងដែរ។ ពីអុកស៊ីហ្សែន នៅក្រោមសកម្មភាពនៃការឆក់អគ្គិសនី អូហ្សូនត្រូវបានបង្កើតឡើង ដែលបង្កើតជាស្រទាប់ការពារអូហ្សូន ដែលការពារជីវិតទាំងអស់នៅលើផែនដីពីផលប៉ះពាល់ដ៏គ្រោះថ្នាក់នៃកាំរស្មីអ៊ុលត្រាវីយូឡេ។
ជនរួមជាតិរបស់យើងដែលជាអ្នកជំនាញខាងសរីរវិទ្យារុក្ខជាតិឆ្នើម K. A. Timiryazev (1843-1920) បានហៅតួនាទីនៃការធ្វើរស្មីសំយោគថា "លោហធាតុ" ចាប់តាំងពីវាភ្ជាប់ផែនដីជាមួយព្រះអាទិត្យ (អវកាស) ផ្តល់លំហូរនៃថាមពលទៅកាន់ភពផែនដី។
ដំណាក់កាលនៃការសំយោគរស្មីសំយោគ។ ប្រតិកម្មពន្លឺនិងងងឹតនៃរស្មីសំយោគទំនាក់ទំនងរបស់ពួកគេ។
នៅឆ្នាំ 1905 អ្នកសរីរវិទ្យារុក្ខជាតិជនជាតិអង់គ្លេស F. Blackman បានរកឃើញថា អត្រានៃការធ្វើរស្មីសំយោគមិនអាចកើនឡើងដោយគ្មានកំណត់ទេ កត្តាមួយចំនួនកំណត់វា។ ដោយផ្អែកលើនេះគាត់បានស្នើឱ្យមានអត្ថិភាពនៃដំណាក់កាលពីរនៃការធ្វើរស្មីសំយោគ: ពន្លឺនិង ងងឹត. នៅអាំងតង់ស៊ីតេពន្លឺទាប ល្បឿននៃប្រតិកម្មពន្លឺកើនឡើងតាមសមាមាត្រទៅនឹងការកើនឡើងនៃអាំងតង់ស៊ីតេពន្លឺ ហើយលើសពីនេះទៀត ប្រតិកម្មទាំងនេះមិនអាស្រ័យលើសីតុណ្ហភាពទេ ព្រោះវាមិនត្រូវការអង់ស៊ីមដើម្បីដំណើរការ។ ប្រតិកម្មពន្លឺកើតឡើងនៅលើភ្នាស thylakoid ។
ផ្ទុយទៅវិញ អត្រានៃប្រតិកម្មងងឹត កើនឡើងជាមួយនឹងការកើនឡើងនៃសីតុណ្ហភាព ទោះជាយ៉ាងណាក៏ដោយ នៅពេលឈានដល់កម្រិតសីតុណ្ហភាព $30°C$ ការលូតលាស់នេះឈប់ ដែលបង្ហាញពីលក្ខណៈអង់ស៊ីមនៃការផ្លាស់ប្តូរដែលកើតឡើងនៅក្នុង stroma ។ វាគួរតែត្រូវបានគេកត់សម្គាល់ថាពន្លឺក៏មានឥទ្ធិពលជាក់លាក់លើប្រតិកម្មងងឹតទោះបីជាការពិតដែលថាពួកគេត្រូវបានគេហៅថាងងឹតក៏ដោយ។

ដំណាក់កាលពន្លឺនៃការធ្វើរស្មីសំយោគដំណើរការលើភ្នាស thylakoid ដែលផ្ទុកនូវប្រភេទប្រូតេអ៊ីនជាច្រើនប្រភេទ ដែលសំខាន់គឺប្រព័ន្ធរូបថត I និង II ក៏ដូចជា ATP synthase ។ សមាសភាពនៃប្រព័ន្ធរូបថតរួមមានសារធាតុពណ៌ ដែលបន្ថែមពីលើក្លរ៉ូហ្វីល ក៏មានសារធាតុ carotenoids ផងដែរ។ Carotenoids ចាប់យកពន្លឺនៅក្នុងតំបន់ទាំងនោះនៃវិសាលគមដែល chlorophyll មិនមាន ហើយក៏ការពារ chlorophyll ពីការបំផ្លាញដោយពន្លឺដែលមានអាំងតង់ស៊ីតេខ្ពស់ផងដែរ។
បន្ថែមពីលើសារធាតុពណ៌ ប្រព័ន្ធរូបថតក៏រួមបញ្ចូលផងដែរនូវប្រូតេអ៊ីនអ្នកទទួលអេឡិចត្រុងមួយចំនួនដែលផ្ទេរអេឡិចត្រុងជាបន្តបន្ទាប់ពីម៉ូលេគុលក្លរ៉ូហ្វីលទៅគ្នាទៅវិញទៅមក។ លំដាប់នៃប្រូតេអ៊ីនទាំងនេះត្រូវបានគេហៅថា ខ្សែសង្វាក់ដឹកជញ្ជូនអេឡិចត្រុង chloroplast ។
ស្មុគស្មាញពិសេសនៃប្រូតេអ៊ីនក៏ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងប្រព័ន្ធ photosystem II ដែលធានានូវការបញ្ចេញអុកស៊ីហ្សែនក្នុងអំឡុងពេលធ្វើរស្មីសំយោគ។ ស្មុគ្រស្មាញដែលវិវត្តន៍ដោយអុកស៊ីហ្សែននេះមានផ្ទុកអ៊ីយ៉ុងម៉ង់ហ្គាណែស និងក្លរីន។
IN ដំណាក់កាលពន្លឺពន្លឺ quanta ឬ photons ដែលធ្លាក់លើម៉ូលេគុល chlorophyll ដែលមានទីតាំងនៅលើភ្នាស thylakoid ផ្ទេរពួកវាទៅស្ថានភាពរំភើបដែលត្រូវបានកំណត់ដោយថាមពលអេឡិចត្រុងខ្ពស់។ ក្នុងពេលជាមួយគ្នានេះដែរ អេឡិចត្រុងដែលរំភើបពី chlorophyll នៃប្រព័ន្ធ photosystem I ត្រូវបានផ្ទេរតាមរយៈខ្សែសង្វាក់អន្តរការីទៅកាន់ក្រុមហ៊ុនដឹកជញ្ជូនអ៊ីដ្រូសែន NADP ដែលបន្ថែមប្រូតុងអ៊ីដ្រូសែន ដែលតែងតែមានវត្តមាននៅក្នុងដំណោះស្រាយ aqueous៖
$NADP + 2e^(-) + 2H^(+) → NADPH + H^(+)$ ។
ការកាត់បន្ថយ $NADPH + H^(+)$ នឹងត្រូវបានប្រើជាបន្តបន្ទាប់ក្នុងដំណាក់កាលងងឹត។ អេឡិចត្រុងពី chlorophyll នៃប្រព័ន្ធ photosystem II ក៏ត្រូវបានផ្ទេរតាមខ្សែសង្វាក់ដឹកជញ្ជូនអេឡិចត្រុងដែរ ប៉ុន្តែពួកគេបានបំពេញ "រន្ធអេឡិចត្រុង" នៅក្នុង chlorophyll នៃប្រព័ន្ធ photosystem I. កង្វះអេឡិចត្រុងនៅក្នុង chlorophyll នៃប្រព័ន្ធ photosystem II ត្រូវបានបំពេញដោយការយកម៉ូលេគុលទឹកចេញពីទឹក។ ម៉ូលេគុលដែលកើតឡើងដោយមានការចូលរួមនៃស្មុគស្មាញបញ្ចេញអុកស៊ីហ្សែនដែលបានរៀបរាប់ខាងលើរួចហើយ។ ជាលទ្ធផលនៃការ decomposition នៃម៉ូលេគុលទឹកដែលត្រូវបានគេហៅថា photolysisអ៊ីដ្រូសែនប្រូតុងត្រូវបានបង្កើតឡើង ហើយអុកស៊ីសែនម៉ូលេគុលត្រូវបានបញ្ចេញ ដែលជាផលិតផលនៃការធ្វើរស្មីសំយោគ៖
$H_2O → 2H^(+) + 2e^(-) + (1)/(2)O_2$ ។
ព័ត៌មានហ្សែននៅក្នុងកោសិកា។ ហ្សែន កូដហ្សែន និងលក្ខណៈសម្បត្តិរបស់វា។ ម៉ាទ្រីសធម្មជាតិនៃប្រតិកម្មជីវសំយោគ។ ជីវសំយោគនៃប្រូតេអ៊ីន និងអាស៊ីត nucleic
ព័ត៌មានហ្សែននៅក្នុងកោសិកា
ការបន្តពូជនៃប្រភេទផ្ទាល់ខ្លួនរបស់មនុស្សម្នាក់គឺជាលក្ខណៈសម្បត្តិជាមូលដ្ឋានមួយនៃការរស់នៅ។ ដោយសារតែបាតុភូតនេះ មានភាពស្រដៀងគ្នាមិនត្រឹមតែរវាងសារពាង្គកាយប៉ុណ្ណោះទេ ថែមទាំងរវាងកោសិកានីមួយៗ ក៏ដូចជាសរីរាង្គរបស់វាផងដែរ (មីតូខនឌ្រី និងផ្លាស្ទីត)។ មូលដ្ឋានសម្ភារៈនៃភាពស្រដៀងគ្នានេះគឺការបញ្ជូនព័ត៌មានហ្សែនដែលត្រូវបានអ៊ិនគ្រីបនៅក្នុងលំដាប់ DNA nucleotide ដែលត្រូវបានអនុវត្តដោយសារតែដំណើរការនៃការចម្លង DNA (ទ្វេដងដោយខ្លួនឯង) ។ លក្ខណៈពិសេស និងលក្ខណៈសម្បត្តិទាំងអស់នៃកោសិកា និងសារពាង្គកាយត្រូវបានដឹងដោយអរគុណចំពោះប្រូតេអ៊ីន រចនាសម្ព័ន្ធដែលត្រូវបានកំណត់ជាចម្បងដោយលំដាប់ DNA nucleotide ។ ដូច្នេះ វាគឺជាការសំយោគជីវសាស្ត្រនៃអាស៊ីត nucleic និងប្រូតេអ៊ីនដែលមានសារៈសំខាន់បំផុតក្នុងដំណើរការមេតាបូលីស។ ឯកតារចនាសម្ព័ន្ធនៃពត៌មានតំណពូជគឺហ្សែន។
ហ្សែន កូដហ្សែន និងលក្ខណៈសម្បត្តិរបស់វា។
ព័ត៌មានតំណពូជនៅក្នុងកោសិកាមួយមិនមែនជា monolithic ទេវាត្រូវបានបែងចែកទៅជា "ពាក្យ" ដាច់ដោយឡែក - ហ្សែន។
ហ្សែនគឺជាឯកតាមូលដ្ឋាននៃព័ត៌មានហ្សែន។
ការងារលើកម្មវិធី "ហ្សែនមនុស្ស" ដែលត្រូវបានអនុវត្តក្នុងពេលដំណាលគ្នាក្នុងប្រទេសមួយចំនួន ហើយត្រូវបានបញ្ចប់នៅដើមសតវត្សនេះ បានផ្តល់ឱ្យយើងនូវការយល់ដឹងថា មនុស្សម្នាក់មានហ្សែនត្រឹមតែ 25-30 ពាន់ហ្សែនប៉ុណ្ណោះ ប៉ុន្តែព័ត៌មានពីភាគច្រើនរបស់យើង DNA មិនត្រូវបានអានទេ ព្រោះវាមានចំនួនច្រើននៃផ្នែកដែលគ្មានន័យ ការនិយាយឡើងវិញ និងការអ៊ិនកូដហ្សែនដែលបាត់បង់អត្ថន័យសម្រាប់មនុស្ស (កន្ទុយ សក់រាងកាយ។ល។)។ លើសពីនេះ ហ្សែនមួយចំនួនដែលទទួលខុសត្រូវចំពោះការវិវត្តនៃជំងឺតំណពូជ ក៏ដូចជាហ្សែនគោលដៅថ្នាំត្រូវបានបកស្រាយ។ ទោះជាយ៉ាងណាក៏ដោយការអនុវត្តជាក់ស្តែងនៃលទ្ធផលដែលទទួលបានក្នុងអំឡុងពេលអនុវត្តកម្មវិធីនេះត្រូវបានពន្យារពេលរហូតដល់ហ្សែនរបស់មនុស្សកាន់តែច្រើនត្រូវបានឌិកូដហើយវាច្បាស់ថាតើវាខុសគ្នាយ៉ាងណា។
ហ្សែនដែលអ៊ិនកូដរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីន ribosomal ឬផ្ទេរ RNA ត្រូវបានគេហៅថា រចនាសម្ព័ន្ធនិងហ្សែនដែលផ្តល់នូវការធ្វើឱ្យសកម្ម ឬការទប់ស្កាត់ការអានព័ត៌មានពីហ្សែនរចនាសម្ព័ន្ធ - បទប្បញ្ញត្តិ. ទោះយ៉ាងណាក៏ដោយ សូម្បីតែហ្សែនរចនាសម្ព័ន្ធក៏មានតំបន់បទប្បញ្ញត្តិដែរ។
ព័ត៌មានតំណពូជនៃសារពាង្គកាយត្រូវបានអ៊ិនគ្រីបនៅក្នុង DNA ក្នុងទម្រង់នៃបន្សំមួយចំនួននៃ nucleotides និងលំដាប់របស់វា - កូដហ្សែន. លក្ខណៈសម្បត្តិរបស់វាគឺ៖ បីដង, ភាពជាក់លាក់, សកល, ភាពលែងត្រូវការតទៅទៀត និងមិនត្រួតស៊ីគ្នា។ លើសពីនេះទៀតមិនមានសញ្ញាវណ្ណយុត្តិនៅក្នុងកូដហ្សែនទេ។
អាស៊ីតអាមីណូនីមួយៗត្រូវបានអ៊ិនកូដនៅក្នុង DNA ដោយនុយក្លេអូទីតបី។ បីដងឧទាហរណ៍ methionine ត្រូវបានអ៊ិនកូដដោយ TAC triplet នោះគឺ លេខកូដ triplet ។ ម៉្យាងវិញទៀត ត្រីភាគីនីមួយៗ អ៊ិនកូដអាស៊ីតអាមីណូតែមួយប៉ុណ្ណោះ ដែលជាភាពជាក់លាក់ ឬភាពមិនច្បាស់លាស់របស់វា។ កូដហ្សែនគឺមានលក្ខណៈជាសកលសម្រាប់សារពាង្គកាយមានជីវិតទាំងអស់ ពោលគឺព័ត៌មានតំណពូជអំពីប្រូតេអ៊ីនរបស់មនុស្សអាចត្រូវបានអានដោយបាក់តេរី និងផ្ទុយមកវិញ។ នេះសក្ខីកម្មដល់ការរួបរួមនៃប្រភពដើមនៃពិភពសរីរាង្គ។ ទោះជាយ៉ាងណាក៏ដោយ 64 បន្សំនៃនុយក្លេអូទីតបីត្រូវគ្នាទៅនឹងអាស៊ីតអាមីណូតែ 20 ដែលជាលទ្ធផលដែល 2-6 បីអាចអ៊ិនកូដអាស៊ីតអាមីណូមួយ នោះគឺជាលេខកូដហ្សែនគឺមិនអាចខ្វះបាន ឬ degenerate ។ កូនបីមិនមានអាស៊ីតអាមីណូដែលត្រូវគ្នាទេ ពួកវាត្រូវបានគេហៅថា បញ្ឈប់ codonsដូចដែលពួកគេសម្គាល់ការបញ្ចប់នៃការសំយោគនៃខ្សែសង្វាក់ polypeptide ។
លំដាប់នៃមូលដ្ឋាននៅក្នុង DNA បីដង និងអាស៊ីតអាមីណូដែលពួកគេបានអ៊ិនកូដ
* បញ្ឈប់ codon ដែលបង្ហាញពីការបញ្ចប់នៃការសំយោគនៃខ្សែសង្វាក់ polypeptide ។
អក្សរកាត់សម្រាប់ឈ្មោះអាស៊ីតអាមីណូ៖
អាឡា - អាឡានីន
Arg - arginine
Asn - asparagine
Asp - អាស៊ីត aspartic
វ៉ាល - វ៉ាលីន
អ៊ីស្ទីឌីន របស់គាត់។
ហ្គីលី - គ្លីសេរីន
Gln - glutamine
Glu - អាស៊ីត glutamic
អ៊ីល - isoleucine
លូ - លូស៊ីន
លីស - លីស៊ីន
ម៉េត - មេតូនីន
ប្រូ - ប្រូលីន
សឺ - សឺរិន
ធីរ - tyrosine
ត្រ - ថូរនីន
បី - tryptophan
ហ្វេន - ហ្វេនីឡាឡាទីន
ស៊ីស្ទីន
ប្រសិនបើអ្នកចាប់ផ្តើមអានព័ត៌មានហ្សែនមិនមែនមកពីនុយក្លេអូទីតទី 1 នៅក្នុង triplet ទេ ប៉ុន្តែពីទីពីរ នោះមិនត្រឹមតែការផ្លាស់ប្តូរស៊ុមនៃការអាននោះទេ - ប្រូតេអ៊ីនដែលត្រូវបានសំយោគតាមរបៀបនេះនឹងខុសគ្នាទាំងស្រុងមិនត្រឹមតែនៅក្នុងលំដាប់នុយក្លេអូទីតប៉ុណ្ណោះទេ ថែមទាំងនៅក្នុងរចនាសម្ព័ន្ធផងដែរ។ និងលក្ខណៈសម្បត្តិ។ មិនមានសញ្ញាវណ្ណយុត្តិរវាងបីទេ ដូច្នេះមិនមានឧបសគ្គចំពោះការផ្លាស់ប្តូរនៃស៊ុមអាន ដែលបើកវិសាលភាពសម្រាប់ការកើតឡើង និងការថែរក្សាការផ្លាស់ប្តូរ។
ម៉ាទ្រីសធម្មជាតិនៃប្រតិកម្មជីវសំយោគ
កោសិកាបាក់តេរីអាចចម្លងរៀងរាល់ 20-30 នាទីម្តង ហើយកោសិកា eukaryotic - ជារៀងរាល់ថ្ងៃ និងច្រើនដងទៀត ដែលទាមទារល្បឿនខ្ពស់ និងភាពត្រឹមត្រូវនៃការចម្លង DNA ។ លើសពីនេះទៀតកោសិកានីមួយៗមានរាប់រយរាប់ពាន់ច្បាប់ចម្លងនៃប្រូតេអ៊ីនជាច្រើនជាពិសេសអង់ស៊ីមដូច្នេះសម្រាប់ការបន្តពូជរបស់ពួកគេវិធីសាស្ត្រ "ដុំ" នៃការផលិតរបស់ពួកគេគឺមិនអាចទទួលយកបានទេ។ មធ្យោបាយរីកចម្រើនជាងនេះគឺការបោះត្រា ដែលអនុញ្ញាតឱ្យអ្នកទទួលបានច្បាប់ចម្លងពិតប្រាកដជាច្រើននៃផលិតផល និងកាត់បន្ថយការចំណាយរបស់វាផងដែរ។ សម្រាប់ការបោះត្រា ត្រូវការម៉ាទ្រីស ដែលការចាប់អារម្មណ៍មួយត្រូវបានធ្វើឡើង។
នៅក្នុងកោសិកាគោលការណ៍នៃការសំយោគម៉ាទ្រីសគឺថាម៉ូលេគុលថ្មីនៃប្រូតេអ៊ីននិងអាស៊ីតនុយក្លេអ៊ីកត្រូវបានសំយោគដោយអនុលោមតាមកម្មវិធីដែលបានដាក់ក្នុងរចនាសម្ព័ន្ធនៃម៉ូលេគុលមុននៃអាស៊ីត nucleic ដូចគ្នា (DNA ឬ RNA) ។
ជីវសំយោគនៃប្រូតេអ៊ីន និងអាស៊ីត nucleic
ការចម្លង DNA ។ DNA គឺជាជីវប៉ូលីម័រដែលមានខ្សែពីរដែលម៉ូណូម័រគឺជានុយក្លេអូទីត។ ប្រសិនបើការសំយោគ DNA ដំណើរការទៅតាមគោលការណ៍នៃការថតចម្លង នោះការបង្ខូចទ្រង់ទ្រាយ និងកំហុសជាច្រើននៅក្នុងព័ត៌មានតំណពូជនឹងកើតឡើងដោយជៀសមិនរួច ដែលនឹងនាំទៅដល់ការស្លាប់របស់សារពាង្គកាយថ្មី។ ដូច្នេះដំណើរការនៃការចម្លង DNA គឺខុសគ្នា។ នៅក្នុងវិធីពាក់កណ្តាលអភិរក្ស៖ ម៉ូលេគុល DNA រំសាយ ហើយនៅលើសង្វាក់នីមួយៗ ខ្សែសង្វាក់ថ្មីត្រូវបានសំយោគដោយយោងទៅតាមគោលការណ៍នៃការបំពេញបន្ថែម។ ដំណើរការនៃការបង្កើតឡើងវិញដោយខ្លួនឯងនៃម៉ូលេគុល DNA ដែលធានានូវការចម្លងពិតប្រាកដនៃព័ត៌មានតំណពូជ និងការបញ្ជូនរបស់វាពីជំនាន់មួយទៅជំនាន់មួយត្រូវបានគេហៅថា ការចម្លង(ពីឡាតាំង។ ការចម្លង- ពាក្យដដែលៗ) ។ ជាលទ្ធផលនៃការចម្លង ច្បាប់ចម្លងពិតប្រាកដចំនួនពីរនៃម៉ូលេគុល DNA មេត្រូវបានបង្កើតឡើង ដែលនីមួយៗមានច្បាប់ចម្លងមួយរបស់មេ។

ដំណើរការនៃការចម្លងគឺពិតជាស្មុគ្រស្មាញខ្លាំងណាស់ ចាប់តាំងពីប្រូតេអ៊ីនមួយចំនួនត្រូវបានចូលរួមនៅក្នុងវា។ ពួកវាខ្លះស្រាយចំណងទ្វេនៃ DNA ខ្លះបំបែកចំណងអ៊ីដ្រូសែនរវាងនុយក្លេអូទីតនៃខ្សែសង្វាក់បំពេញបន្ថែម ខ្លះទៀត (ឧទាហរណ៍ អង់ស៊ីម DNA polymerase) ជ្រើសរើសនុយក្លេអូទីតថ្មីតាមគោលការណ៍នៃការបំពេញបន្ថែម។ល។ ម៉ូលេគុល DNA ពីរបានបង្កើតឡើងជាលទ្ធផលនៃ កោសិកាកូនស្រីដែលទើបបង្កើតថ្មី
កំហុសក្នុងដំណើរការចម្លងគឺកម្រមានណាស់ ប៉ុន្តែប្រសិនបើវាកើតឡើង ពួកគេត្រូវបានលុបចោលយ៉ាងឆាប់រហ័សដោយទាំង DNA polymerases និងអង់ស៊ីមជួសជុលពិសេស ចាប់តាំងពីកំហុសណាមួយនៅក្នុងលំដាប់នុយក្លេអូទីតអាចនាំឱ្យមានការផ្លាស់ប្តូរដែលមិនអាចត្រឡប់វិញបាននៅក្នុងរចនាសម្ព័ន្ធ និងមុខងារនៃប្រូតេអ៊ីន។ ហើយនៅទីបំផុត ប៉ះពាល់យ៉ាងធ្ងន់ធ្ងរដល់លទ្ធភាពជោគជ័យនៃកោសិកាថ្មី ឬសូម្បីតែបុគ្គល។
ជីវសំយោគប្រូតេអ៊ីន។ដូចដែលទស្សនវិទូឆ្នើមនៃសតវត្សទី 19 F. Engels បានដាក់ន័យធៀបវាថា "ជីវិតគឺជាទម្រង់នៃអត្ថិភាពនៃសាកសពប្រូតេអ៊ីន" ។ រចនាសម្ព័ន្ធ និងលក្ខណៈសម្បត្តិនៃម៉ូលេគុលប្រូតេអ៊ីនត្រូវបានកំណត់ដោយរចនាសម្ព័ន្ធចម្បងរបស់វា ពោលគឺ លំដាប់នៃអាស៊ីតអាមីណូដែលបានអ៊ិនកូដនៅក្នុង DNA ។ មិនត្រឹមតែអត្ថិភាពនៃសារធាតុ polypeptide ខ្លួនវាប៉ុណ្ណោះទេ ប៉ុន្តែវាក៏មានដំណើរការនៃកោសិកាទាំងមូលផងដែរ អាស្រ័យលើភាពត្រឹមត្រូវនៃការបន្តពូជនៃព័ត៌មាននេះ ដូច្នេះហើយ ដំណើរការនៃការសំយោគប្រូតេអ៊ីនមានសារៈសំខាន់ណាស់។ វាហាក់ដូចជាដំណើរការស្មុគស្មាញបំផុតនៃការសំយោគនៅក្នុងកោសិកា ចាប់តាំងពីអង់ស៊ីមរហូតដល់ទៅបីរយផ្សេងគ្នា និងម៉ាក្រូម៉ូលេគុលផ្សេងទៀតត្រូវបានចូលរួមនៅទីនេះ។ លើសពីនេះទៀតវាហូរក្នុងល្បឿនលឿនដែលទាមទារភាពជាក់លាក់កាន់តែច្រើន។
មានជំហានសំខាន់ពីរក្នុងការសំយោគប្រូតេអ៊ីន៖ ការចម្លង និងការបកប្រែ។
ប្រតិចារិក(ពីឡាតាំង។ ប្រតិចារិក- សរសេរឡើងវិញ) គឺជាការសំយោគជីវសាស្ត្រនៃម៉ូលេគុល mRNA នៅលើគំរូ DNA ។

ដោយសារម៉ូលេគុល DNA មានច្រវាក់ប្រឆាំងប៉ារ៉ាឡែលពីរ ការអានព័ត៌មានពីខ្សែសង្វាក់ទាំងពីរនឹងនាំទៅរកការបង្កើត mRNAs ខុសគ្នាទាំងស្រុង ដូច្នេះការសំយោគជីវសាស្ត្ររបស់ពួកវាគឺអាចធ្វើទៅបានតែលើខ្សែសង្វាក់មួយប៉ុណ្ណោះ ដែលត្រូវបានគេហៅថា coding ឬ codogenic ផ្ទុយទៅនឹងទីពីរ។ មិនសរសេរកូដ ឬមិនមែន codogenic ។ ដំណើរការសរសេរឡើងវិញត្រូវបានផ្តល់ដោយអង់ស៊ីមពិសេស RNA polymerase ដែលជ្រើសរើស RNA nucleotides តាមគោលការណ៍នៃការបំពេញបន្ថែម។ ដំណើរការនេះអាចកើតឡើងទាំងនៅក្នុងស្នូល និងនៅក្នុងសរីរាង្គដែលមាន DNA ផ្ទាល់របស់ពួកគេ - mitochondria និង plastids ។
ម៉ូលេគុល mRNA ដែលត្រូវបានសំយោគកំឡុងពេលចម្លងឆ្លងកាត់ដំណើរការស្មុគស្មាញនៃការរៀបចំសម្រាប់ការបកប្រែ (mRNAs mitochondrial និង plastid អាចស្ថិតនៅក្នុងសរីរាង្គ ដែលដំណាក់កាលទីពីរនៃការសំយោគប្រូតេអ៊ីនកើតឡើង)។ នៅក្នុងដំណើរការនៃភាពចាស់ទុំរបស់ mRNA នុយក្លេអូទីតបីដំបូង (AUG) និងកន្ទុយនៃ adenyl nucleotides ត្រូវបានភ្ជាប់ទៅវា ដែលប្រវែងកំណត់ថាតើអាចសំយោគប្រូតេអ៊ីនប៉ុន្មាននៅលើម៉ូលេគុលដែលបានផ្តល់ឱ្យ។ មានតែពេលនោះទេដែល mRNAs ចាស់ទុំចាកចេញពីស្នូលតាមរយៈរន្ធនុយក្លេអ៊ែរ។
ស្របគ្នាដំណើរការនៃការធ្វើឱ្យអាស៊ីតអាមីណូកើតឡើងនៅក្នុង cytoplasm ក្នុងអំឡុងពេលដែលអាស៊ីតអាមីណូត្រូវបានភ្ជាប់ទៅនឹង tRNA ឥតគិតថ្លៃដែលត្រូវគ្នា។ ដំណើរការនេះត្រូវបានជំរុញដោយអង់ស៊ីមពិសេសវាស៊ី ATP ។
ផ្សាយ(ពីឡាតាំង។ ការចាក់ផ្សាយ- ការផ្ទេរ) គឺជាការសំយោគជីវសាស្ត្រនៃសង្វាក់ polypeptide នៅលើគំរូ mRNA ដែលក្នុងនោះព័ត៌មានហ្សែនត្រូវបានបកប្រែទៅជាលំដាប់នៃអាស៊ីតអាមីណូនៃខ្សែសង្វាក់ polypeptide ។

ដំណាក់កាលទីពីរនៃការសំយោគប្រូតេអ៊ីនច្រើនតែកើតឡើងនៅក្នុង cytoplasm ឧទាហរណ៍នៅលើរដុប endoplasmic reticulum ។ ការកើតឡើងរបស់វាតម្រូវឱ្យមានវត្តមានរបស់ ribosomes ការធ្វើឱ្យសកម្មនៃ tRNA ក្នុងអំឡុងពេលដែលពួកវាភ្ជាប់អាស៊ីតអាមីណូដែលត្រូវគ្នា វត្តមាននៃអ៊ីយ៉ុង Mg2+ ក៏ដូចជាលក្ខខណ្ឌបរិស្ថានដ៏ល្អប្រសើរ (សីតុណ្ហភាព pH សម្ពាធជាដើម)។
ដើម្បីចាប់ផ្តើមការចាក់ផ្សាយ ការចាប់ផ្តើម) ផ្នែកតូចមួយនៃ ribosome ត្រូវបានភ្ជាប់ទៅនឹងម៉ូលេគុល mRNA ដែលត្រៀមរួចជាស្រេចសម្រាប់ការសំយោគ ហើយបន្ទាប់មកយោងទៅតាមគោលការណ៍នៃការបំពេញបន្ថែម tRNA ដែលផ្ទុកអាស៊ីតអាមីណូ methionine ត្រូវបានជ្រើសរើសទៅ codon ដំបូង (AUG) ។ មានតែពេលនោះទេដែលអនុផ្នែកធំនៃ ribosome ចូលរួម។ នៅក្នុង ribosome ដែលបានជួបប្រជុំគ្នា មាន mRNA codons ពីរ ដែលទីមួយត្រូវបានកាន់កាប់រួចហើយ។ tRNA ទីពីរដែលផ្ទុកអាស៊ីតអាមីណូផងដែរត្រូវបានភ្ជាប់ទៅនឹង codon ដែលនៅជាប់នឹងវាបន្ទាប់មកចំណង peptide ត្រូវបានបង្កើតឡើងរវាងសំណល់អាស៊ីតអាមីណូដោយមានជំនួយពីអង់ស៊ីម។ ribosome ផ្លាស់ទី codon មួយនៃ mRNA; ទីមួយនៃ tRNA ដែលត្រូវបានដោះលែងពីអាស៊ីតអាមីណូ ត្រឡប់ទៅស៊ីតូប្លាសសម្រាប់អាស៊ីតអាមីណូបន្ទាប់ ហើយបំណែកនៃខ្សែសង្វាក់ polypeptide នាពេលអនាគតដូចដែលវាមាន ព្យួរនៅលើ tRNA ដែលនៅសល់។ tRNA បន្ទាប់ចូលរួមជាមួយ codon ថ្មី ដែលស្ថិតនៅក្នុង ribosome ដំណើរការនេះកើតឡើងវិញ ហើយជាជំហានៗ ខ្សែសង្វាក់ polypeptide លាតសន្ធឹង ពោលគឺវា ការពន្លូត។
ចុងបញ្ចប់នៃការសំយោគប្រូតេអ៊ីន ការបញ្ចប់) កើតឡើងភ្លាមៗនៅពេលដែលលំដាប់នុយក្លេអូទីតជាក់លាក់មួយត្រូវបានជួបប្រទះនៅក្នុងម៉ូលេគុល mRNA ដែលមិនបានអ៊ិនកូដអាស៊ីតអាមីណូ (បញ្ឈប់ codon) ។ បន្ទាប់ពីនោះ ribosome, mRNA និងខ្សែសង្វាក់ polypeptide ត្រូវបានបំបែកចេញ ហើយប្រូតេអ៊ីនដែលបានសំយោគថ្មីទទួលបានរចនាសម្ព័ន្ធសមស្រប ហើយត្រូវបានបញ្ជូនទៅផ្នែកនៃកោសិកាដែលវានឹងបំពេញមុខងាររបស់វា។
ការបកប្រែគឺជាដំណើរការដែលប្រើប្រាស់ថាមពលខ្លាំង ចាប់តាំងពីថាមពលនៃម៉ូលេគុល ATP មួយត្រូវបានចំណាយលើការភ្ជាប់អាស៊ីតអាមីណូមួយទៅ tRNA ហើយមួយចំនួនទៀតត្រូវបានប្រើដើម្បីផ្លាស់ទី ribosome តាមម៉ូលេគុល mRNA ។
ដើម្បីពន្លឿនការសំយោគនៃម៉ូលេគុលប្រូតេអ៊ីនមួយចំនួន ribosomes ជាច្រើនអាចត្រូវបានភ្ជាប់ជាបន្តបន្ទាប់ទៅនឹងម៉ូលេគុល mRNA ដែលបង្កើតបានជារចនាសម្ព័ន្ធតែមួយ - ប៉ូលីសូម។
កោសិកាគឺជាឯកតាហ្សែននៃភាវៈរស់។ ក្រូម៉ូសូមរចនាសម្ព័ន្ធ (រូបរាង និងទំហំ) និងមុខងារ។ ចំនួនក្រូម៉ូសូម និងប្រភេទរបស់វាថេរ។ កោសិកា Somatic និងផ្លូវភេទ។ វដ្តជីវិតកោសិកា៖ អន្តរដំណាក់កាល និង មីតូស៊ីស។ Mitosis គឺជាការបែងចែកកោសិកា somatic ។ ម៉ីអូស៊ីស។ ដំណាក់កាលនៃ mitosis និង meiosis ។ ការវិវត្តន៍នៃកោសិកាមេរោគនៅក្នុងរុក្ខជាតិ និងសត្វ។ ការបែងចែកកោសិកាគឺជាមូលដ្ឋានសម្រាប់ការលូតលាស់ ការអភិវឌ្ឍន៍ និងការបន្តពូជរបស់សារពាង្គកាយ។ តួនាទីរបស់ meiosis និង mitosis
កោសិកាគឺជាអង្គភាពហ្សែននៃជីវិត
ទោះបីជាការពិតដែលថាអាស៊ីត nucleic គឺជាអ្នកបញ្ជូនព័ត៌មានហ្សែនក៏ដោយការសម្រេចនៃព័ត៌មាននេះគឺមិនអាចទៅរួចទេនៅខាងក្រៅកោសិកាដែលត្រូវបានបង្ហាញយ៉ាងងាយស្រួលដោយឧទាហរណ៍នៃមេរោគ។ សារពាង្គកាយទាំងនេះ ច្រើនតែមាន DNA ឬ RNA មិនអាចបង្កើតឡើងវិញដោយខ្លួនឯងបានទេ សម្រាប់ការនេះ ពួកគេត្រូវតែប្រើឧបករណ៍តំណពូជនៃកោសិកា។ ពួកវាមិនអាចសូម្បីតែជ្រាបចូលទៅក្នុងកោសិកាដោយគ្មានជំនួយពីកោសិកាដោយខ្លួនឯង លើកលែងតែតាមរយៈការប្រើប្រាស់យន្តការដឹកជញ្ជូនភ្នាស ឬដោយសារការខូចខាតកោសិកា។ មេរោគភាគច្រើនមិនស្ថិតស្ថេរ ពួកវាស្លាប់បន្ទាប់ពីការប៉ះពាល់នឹងខ្យល់អាកាសពីរបីម៉ោង។ ដូច្នេះកោសិកាគឺជាឯកតាហ្សែននៃការរស់នៅដែលមានសំណុំអប្បបរមានៃសមាសធាតុសម្រាប់ការអភិរក្ស ការកែប្រែ និងការអនុវត្តព័ត៌មានតំណពូជ ក៏ដូចជាការបញ្ជូនរបស់វាទៅកូនចៅ។
ព័ត៌មានហ្សែនភាគច្រើននៃកោសិកា eukaryotic មានទីតាំងនៅស្នូល។ លក្ខណៈពិសេសនៃអង្គការរបស់វាគឺថាមិនដូច DNA នៃកោសិកា prokaryotic ទេម៉ូលេគុល DNA eukaryotic មិនត្រូវបានបិទហើយបង្កើតជាស្មុគស្មាញស្មុគស្មាញជាមួយប្រូតេអ៊ីន - ក្រូម៉ូសូម។
ក្រូម៉ូសូមរចនាសម្ព័ន្ធ (រូបរាង និងទំហំ) និងមុខងារ
ក្រូម៉ូសូម(មកពីភាសាក្រិក។ ក្រូម- ពណ៌ពណ៌និង ត្រីឆ្មា- រាងកាយ) គឺជារចនាសម្ព័ន្ធនៃស្នូលកោសិកាដែលមានហ្សែន និងផ្ទុកព័ត៌មានតំណពូជជាក់លាក់អំពីសញ្ញា និងលក្ខណៈសម្បត្តិនៃរាងកាយ។
ពេលខ្លះ ម៉ូលេគុល DNA របស់ prokaryotes ត្រូវបានគេហៅថា ក្រូម៉ូសូមផងដែរ។ ក្រូម៉ូសូមមានសមត្ថភាពចម្លងដោយខ្លួនឯង ពួកគេមានរចនាសម្ព័ន្ធ និងមុខងារបុគ្គល ហើយរក្សាវាទុកក្នុងជំនាន់ជាច្រើន។ កោសិកានីមួយៗផ្ទុកព័ត៌មានតំណពូជទាំងអស់នៃរាងកាយ ប៉ុន្តែមានតែផ្នែកតូចមួយប៉ុណ្ណោះដែលវាដំណើរការ។
មូលដ្ឋាននៃក្រូម៉ូសូមគឺជាម៉ូលេគុល DNA ពីរខ្សែដែលផ្ទុកដោយប្រូតេអ៊ីន។ នៅក្នុង eukaryotes ប្រូតេអ៊ីន histone និង non-histone ធ្វើអន្តរកម្មជាមួយ DNA ខណៈពេលដែលនៅក្នុង prokaryotes ប្រូតេអ៊ីន histone គឺអវត្តមាន។
ក្រូម៉ូសូមត្រូវបានគេមើលឃើញល្អបំផុតនៅក្រោមមីក្រូទស្សន៍ពន្លឺកំឡុងពេលបែងចែកកោសិកា នៅពេលដែលជាលទ្ធផលនៃការបង្រួម ពួកវាយកទម្រង់ជារាងជាដំបងដែលបំបែកដោយការបង្រួមបឋម - កណ្តាល — នៅលើស្មា. ក្រូម៉ូសូមក៏អាចមានដែរ។ ការរឹតបន្តឹងបន្ទាប់បន្សំដែលក្នុងករណីខ្លះបំបែកអ្វីដែលគេហៅថា ផ្កាយរណប. ចុងបញ្ចប់នៃក្រូម៉ូសូមត្រូវបានគេហៅថា តេឡូមេស. Telomeres រារាំងចុងបញ្ចប់នៃក្រូម៉ូសូមមិនឱ្យនៅជាប់គ្នា និងធានាការភ្ជាប់របស់វាទៅនឹងភ្នាសនុយក្លេអ៊ែរនៅក្នុងកោសិកាដែលមិនបែងចែក។ នៅដើមដំបូងនៃការបែងចែក ក្រូម៉ូសូមត្រូវបានកើនឡើងទ្វេដង និងមានក្រូម៉ូសូមកូនស្រីពីរ - ក្រូម៉ាទីតភ្ជាប់នៅកណ្តាល។

យោងតាមរូបរាង ក្រូម៉ូសូមដៃស្មើគ្នា ដៃមិនស្មើគ្នា និងរាងជាដំបងត្រូវបានសម្គាល់។ ទំហំក្រូម៉ូសូមប្រែប្រួលយ៉ាងខ្លាំង ប៉ុន្តែក្រូម៉ូសូមមធ្យមមានទំហំ 5$×$1.4 µm។
ក្នុងករណីខ្លះ ក្រូម៉ូសូមដែលជាលទ្ធផលនៃការចម្លង DNA ជាច្រើនមានក្រូម៉ាទីតរាប់រយរាប់ពាន់៖ ក្រូម៉ូសូមយក្សបែបនេះត្រូវបានគេហៅថា ប៉ូលីថេន. ពួកវាត្រូវបានរកឃើញនៅក្នុងក្រពេញទឹកមាត់របស់ដង្កូវ Drosophila ក៏ដូចជានៅក្នុងក្រពេញរំលាយអាហាររបស់ដង្កូវមូល។
ចំនួនក្រូម៉ូសូម និងប្រភេទរបស់វាថេរ។ កោសិកា Somatic និងមេរោគ
យោងតាមទ្រឹស្ដីកោសិកា កោសិកាគឺជាឯកតានៃរចនាសម្ព័ន្ធ ជីវិត និងការអភិវឌ្ឍន៍នៃសារពាង្គកាយ។ ដូច្នេះមុខងារសំខាន់ៗរបស់ភាវៈមានជីវិតដូចជាការលូតលាស់ ការបន្តពូជ និងការអភិវឌ្ឍន៍នៃសារពាង្គកាយត្រូវបានផ្តល់នៅកម្រិតកោសិកា។ កោសិកានៃសារពាង្គកាយពហុកោសិកាអាចត្រូវបានបែងចែកទៅជា somatic និងភេទ។
កោសិកា somaticគឺជាកោសិកាទាំងអស់នៃរាងកាយដែលត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែក mitotic ។
ការសិក្សាអំពីក្រូម៉ូសូមបានធ្វើឱ្យវាអាចធ្វើទៅបានដើម្បីកំណត់ថាកោសិកា somatic នៃសារពាង្គកាយនៃប្រភេទជីវសាស្រ្តនីមួយៗត្រូវបានកំណត់លក្ខណៈដោយចំនួនក្រូម៉ូសូមថេរ។ ឧទាហរណ៍ មនុស្សម្នាក់មាន 46 ពួកវា។ សំណុំនៃក្រូម៉ូសូមនៃកោសិកា somatic ត្រូវបានគេហៅថា diploid(2n) ឬទ្វេដង។

កោសិកាផ្លូវភេទ, ឬ gametesគឺជាកោសិកាពិសេសដែលបម្រើសម្រាប់ការបន្តពូជផ្លូវភេទ។
Gametes តែងតែមានក្រូម៉ូសូមពាក់កណ្តាលជាច្រើនដូចជាកោសិកា somatic (នៅក្នុងមនុស្ស - 23) ដូច្នេះសំណុំនៃក្រូម៉ូសូមនៃកោសិកាមេរោគត្រូវបានគេហៅថា haploid(n) ឬនៅលីវ។ ការបង្កើតរបស់វាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបែងចែកកោសិកា meiotic ។
បរិមាណ DNA នៃកោសិកា somatic ត្រូវបានកំណត់ថាជា 2c ហើយកោសិកាមេរោគគឺ 1c ។ រូបមន្តហ្សែននៃកោសិកា somatic ត្រូវបានសរសេរជា 2n2c និងភេទ - 1n1c ។
នៅក្នុងស្នូលនៃកោសិកា somatic មួយចំនួន ចំនួននៃក្រូម៉ូសូមអាចខុសគ្នាពីចំនួនរបស់ពួកគេនៅក្នុងកោសិកា somatic ។ ប្រសិនបើភាពខុសគ្នានេះធំជាងដោយមួយ ពីរ បី ជាដើម។ សំណុំ haploid នោះកោសិកាបែបនេះត្រូវបានគេហៅថា polyploid(tri-, tetra-, pentaploid រៀងគ្នា) ។ នៅក្នុងកោសិកាបែបនេះ ដំណើរការមេតាបូលីសជាធម្មតាពឹងផ្អែកខ្លាំង។
ចំនួនក្រូម៉ូសូមនៅក្នុងខ្លួនវាមិនមែនជាលក្ខណៈជាក់លាក់នៃប្រភេទសត្វទេ ដោយសារសារពាង្គកាយផ្សេងៗគ្នាអាចមានចំនួនក្រូម៉ូសូមដូចគ្នា ខណៈដែលសារធាតុដែលពាក់ព័ន្ធអាចមានលេខខុសៗគ្នា។ ជាឧទាហរណ៍ ជំងឺគ្រុនចាញ់ plasmodium និងដង្កូវមូលសេះមានក្រូម៉ូសូមពីរ ខណៈមនុស្ស និងសត្វស្វាមាន 46 និង 48 រៀងគ្នា។
ក្រូម៉ូសូមរបស់មនុស្សត្រូវបានបែងចែកជាពីរក្រុម៖ អូតូសូម និងក្រូម៉ូសូមភេទ (heterochromosomes) ។ ស្វ័យភាពមាន 22 គូនៅក្នុងកោសិកា somatic របស់មនុស្ស, ពួកគេគឺដូចគ្នាសម្រាប់បុរសនិងស្ត្រី, និង ក្រូម៉ូសូមផ្លូវភេទមួយគូ ប៉ុន្តែវាគឺជានាងដែលកំណត់ភេទរបស់បុគ្គល។ ក្រូម៉ូសូមផ្លូវភេទមានពីរប្រភេទគឺ X និង Y. កោសិកានៃរាងកាយរបស់ស្ត្រីផ្ទុកក្រូម៉ូសូម X ពីរហើយបុរស - X និង Y ។
Karyotype- នេះគឺជាសំណុំនៃសញ្ញានៃសំណុំក្រូម៉ូសូមនៃសារពាង្គកាយមួយ (ចំនួនក្រូម៉ូសូម រូបរាង និងទំហំរបស់វា)។
កំណត់ត្រាតាមលក្ខខណ្ឌនៃ karyotype រួមមានចំនួនសរុបនៃក្រូម៉ូសូម ក្រូម៉ូសូមភេទ និងគម្លាតដែលអាចកើតមាននៅក្នុងសំណុំនៃក្រូម៉ូសូម។ ឧទាហរណ៍ karyotype នៃបុរសធម្មតាត្រូវបានសរសេរជា 46,XY ខណៈពេលដែល karyotype របស់ស្ត្រីធម្មតាគឺ 46,XX ។
វដ្តជីវិតកោសិកា៖ អន្តរដំណាក់កាល និង មីតូស៊ីស
កោសិកាមិនកើតឡើងរាល់ពេលថ្មីទេ វាត្រូវបានបង្កើតឡើងតែជាលទ្ធផលនៃការបែងចែកកោសិកាម្តាយប៉ុណ្ណោះ។ បន្ទាប់ពីការបែកគ្នា កោសិកាកូនស្រីត្រូវចំណាយពេលខ្លះដើម្បីបង្កើតសរីរាង្គ និងទទួលបានរចនាសម្ព័ន្ធសមស្រប ដែលនឹងធានាបាននូវការអនុវត្តមុខងារជាក់លាក់មួយ។ រយៈពេលនេះត្រូវបានគេហៅថា ការទុំ។
រយៈពេលនៃការលេចឡើងនៃកោសិកាដែលជាលទ្ធផលនៃការបែងចែកទៅជាការបែងចែកឬការស្លាប់របស់វាត្រូវបានគេហៅថា វដ្តជីវិតកោសិកា។
នៅក្នុងកោសិកា eukaryotic វដ្តជីវិតត្រូវបានបែងចែកជាពីរដំណាក់កាលសំខាន់គឺ interphase និង mitosis ។
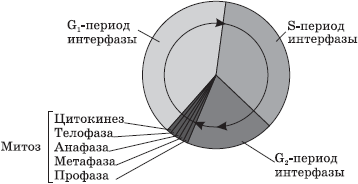
អន្តរដំណាក់កាល- នេះគឺជារយៈពេលនៃវដ្តជីវិតដែលកោសិកាមិនបែងចែក និងដំណើរការជាធម្មតា។ interphase ត្រូវបានបែងចែកជាបីដំណាក់កាល៖ G 1 -, S- និង G 2 -periods ។
G 1 - រយៈពេល(presynthetic, postmitotic) គឺជាកំឡុងពេលនៃការលូតលាស់ និងការអភិវឌ្ឍន៍កោសិកា ដែលកំឡុងពេលនោះមានការសំយោគសកម្មនៃ RNA ប្រូតេអ៊ីន និងសារធាតុផ្សេងៗទៀតដែលចាំបាច់សម្រាប់ជំនួយជីវិតពេញលេញនៃកោសិកាដែលទើបបង្កើតថ្មី។ នៅចុងបញ្ចប់នៃរយៈពេលនេះ កោសិកាអាចចាប់ផ្តើមរៀបចំសម្រាប់ការចម្លង DNA ។
IN S-period(សំយោគ) ដំណើរការនៃការចម្លង DNA កើតឡើង។ ផ្នែកតែមួយគត់នៃក្រូម៉ូសូមដែលមិនឆ្លងកាត់ការចម្លងគឺ centromere ដូច្នេះម៉ូលេគុល DNA លទ្ធផលមិនខុសគ្នាទាំងស្រុងនោះទេ ប៉ុន្តែនៅតែជាប់នៅក្នុងវា ហើយនៅដើមដំបូងនៃការបែងចែក ក្រូម៉ូសូមមានរូបរាងរាងអក្សរ X។ រូបមន្តហ្សែននៃកោសិកាបន្ទាប់ពីការចម្លង DNA គឺ 2n4c ។ ផងដែរនៅក្នុង S-period ការកើនឡើងទ្វេដងនៃ centrioles នៃមជ្ឈមណ្ឌលកោសិកាកើតឡើង។
G 2 - រយៈពេល(postsynthetic, premitotic) ត្រូវបានកំណត់ដោយការសំយោគដែលពឹងផ្អែកខ្លាំងនៃ RNA ប្រូតេអ៊ីននិង ATP ដែលចាំបាច់សម្រាប់ដំណើរការនៃការបែងចែកកោសិកាក៏ដូចជាការបំបែកនៃ centrioles, mitochondria និង plastids ។ រហូតដល់ចុងបញ្ចប់នៃ interphase, chromatin និង nucleolus នៅតែអាចបែងចែកបានយ៉ាងច្បាស់ ភាពសុចរិតនៃភ្នាសនុយក្លេអ៊ែរមិនត្រូវបានរំខាន ហើយសរីរាង្គមិនផ្លាស់ប្តូរទេ។
កោសិកាមួយចំនួននៃរាងកាយអាចបំពេញមុខងាររបស់ពួកគេពេញមួយជីវិតរបស់រាងកាយ (ណឺរ៉ូននៃខួរក្បាលរបស់យើង កោសិកាសាច់ដុំនៃបេះដូង) ខណៈពេលដែលកោសិកាខ្លះទៀតមានក្នុងរយៈពេលខ្លី ក្រោយមកពួកគេស្លាប់ (កោសិកានៃ epithelium ពោះវៀន។ , កោសិកានៃ epidermis នៃស្បែក) ។ អាស្រ័យហេតុនេះ ដំណើរការនៃការបែងចែកកោសិកា និងការបង្កើតកោសិកាថ្មីត្រូវតែកើតឡើងជានិច្ចនៅក្នុងខ្លួន ដែលនឹងជំនួសកោសិកាដែលស្លាប់។ ក្រឡាដែលមានសមត្ថភាពបែងចែកត្រូវបានគេហៅថា ដើម. នៅក្នុងខ្លួនមនុស្ស ពួកវាត្រូវបានរកឃើញនៅក្នុងខួរឆ្អឹងក្រហម នៅក្នុងស្រទាប់ជ្រៅនៃ epidermis នៃស្បែក និងកន្លែងផ្សេងៗទៀត។ ដោយប្រើកោសិកាទាំងនេះ អ្នកអាចបណ្តុះសរីរាង្គថ្មី សម្រេចបាននូវភាពស្រស់ថ្លា និងក្លូនរាងកាយផងដែរ។ ការរំពឹងទុកសម្រាប់ការប្រើប្រាស់កោសិកាដើមគឺមានភាពច្បាស់លាស់ ប៉ុន្តែទិដ្ឋភាពសីលធម៌ និងសីលធម៌នៃបញ្ហានេះនៅតែត្រូវបានពិភាក្សានៅឡើយ ចាប់តាំងពីក្នុងករណីភាគច្រើន កោសិកាដើមអំប្រ៊ីយ៉ុងដែលទទួលបានពីទារកដែលបានស្លាប់ក្នុងអំឡុងពេលរំលូតកូនត្រូវបានប្រើ។
រយៈពេលនៃ interphase នៅក្នុងកោសិការុក្ខជាតិនិងសត្វជាមធ្យម 10-20 ម៉ោងខណៈពេលដែល mitosis ចំណាយពេលប្រហែល 1-2 ម៉ោង។
ក្នុងអំឡុងពេលនៃការបែងចែកជាបន្តបន្ទាប់នៅក្នុងសារពាង្គកាយពហុកោសិកា កោសិកាកូនស្រីកាន់តែមានភាពចម្រុះ ចាប់តាំងពីពួកគេបានអានព័ត៌មានពីចំនួនហ្សែនកើនឡើង។
កោសិកាមួយចំនួននៅទីបំផុតបញ្ឈប់ការបែងចែក និងស្លាប់ ដែលអាចបណ្តាលមកពីការបំពេញមុខងារជាក់លាក់ ដូចជានៅក្នុងករណីនៃកោសិកាស្បែក និងកោសិកាឈាម ឬបំផ្លាញកោសិកាទាំងនេះដោយកត្តាបរិស្ថាន ជាពិសេសភ្នាក់ងារបង្កជំងឺ។ ការស្លាប់កោសិកាតាមកម្មវិធីហ្សែនត្រូវបានគេហៅថា apoptosisខណៈពេលដែលការស្លាប់ដោយចៃដន្យ necrosis.
Mitosis គឺជាការបែងចែកកោសិកា somatic ។ ដំណាក់កាលនៃ mitosis
មីតូស៊ីស- វិធីសាស្រ្តនៃការបែងចែកដោយប្រយោលនៃកោសិកា somatic ។
ក្នុងអំឡុងពេល mitosis កោសិកាឆ្លងកាត់ដំណាក់កាលបន្តបន្ទាប់គ្នា ជាលទ្ធផលដែលកោសិកាកូនស្រីនីមួយៗទទួលបានសំណុំក្រូម៉ូសូមដូចគ្នាដូចនៅក្នុងកោសិកាម្តាយ។
Mitosis ត្រូវបានបែងចែកជា 4 ដំណាក់កាលសំខាន់ៗគឺ prophase, metaphase, anaphase និង telophase ។ ប្រូហ្វាស- ដំណាក់កាលដ៏វែងបំផុតនៃ mitosis ក្នុងអំឡុងពេលដែលការ condensation ក្រូម៉ូសូមកើតឡើង ជាលទ្ធផលដែលក្រូម៉ូសូមរាង X ដែលមានក្រូម៉ាទីតពីរ (ក្រូម៉ូសូមកូនស្រី) អាចមើលឃើញ។ ក្នុងករណីនេះ nucleolus រលាយបាត់ centrioles បង្វែរទៅរកប៉ូលនៃកោសិកា ហើយ achromatin spindle (spindle) នៃ microtubules ចាប់ផ្តើមបង្កើត។ នៅចុងបញ្ចប់នៃ prophase ភ្នាសនុយក្លេអ៊ែរបំបែកទៅជា vesicles ដាច់ដោយឡែក។

IN metaphaseក្រូម៉ូសូមតម្រង់ជួរតាមបណ្តោយខ្សែអេក្វាទ័រនៃកោសិកាជាមួយនឹង centromeres របស់ពួកគេ ដែល microtubules នៃ spindle បែងចែកដែលបានបង្កើតឡើងយ៉ាងពេញលេញត្រូវបានភ្ជាប់។ នៅដំណាក់កាលនៃការបែងចែកនេះ ក្រូម៉ូសូមគឺក្រាស់បំផុត និងមានរូបរាងលក្ខណៈ ដែលធ្វើឱ្យវាអាចសិក្សាពី karyotype ។
IN អាណាហ្វាសការចម្លង DNA យ៉ាងឆាប់រហ័សកើតឡើងនៅក្នុង centromeres ដែលជាលទ្ធផលដែលក្រូម៉ូសូមបំបែក ហើយក្រូម៉ាទីតបង្វែរទៅប៉ូលនៃកោសិកា ដែលលាតសន្ធឹងដោយ microtubules ។ ការចែកចាយក្រូម៉ាទីតត្រូវតែស្មើគ្នាទាំងស្រុង ព្រោះវាជាដំណើរការនេះដែលរក្សាភាពថេរនៃចំនួនក្រូម៉ូសូមនៅក្នុងកោសិកានៃរាងកាយ។
នៅលើឆាក តេឡូហ្វាសក្រូម៉ូសូមរបស់កូនស្រីប្រមូលផ្តុំនៅប៉ូល ធ្វើឱ្យខូចចិត្ត ជុំវិញស្រោមសំបុត្រនុយក្លេអ៊ែរបង្កើតចេញពី vesicles ហើយ nucleoli លេចឡើងនៅក្នុង nuclei ដែលទើបបង្កើតថ្មី។
បន្ទាប់ពីការបែងចែកស្នូល ការបែងចែកស៊ីតូប្លាសកើតឡើង - cytokinesis,ក្នុងអំឡុងពេលនោះមានការចែកចាយឯកសណ្ឋានច្រើនឬតិចនៃសរីរាង្គទាំងអស់នៃកោសិកាមេ។
ដូច្នេះជាលទ្ធផលនៃ mitosis កោសិកាកូនស្រីពីរត្រូវបានបង្កើតឡើងពីកោសិកាម្តាយមួយ ដែលកោសិកានីមួយៗគឺជាច្បាប់ចម្លងហ្សែននៃកោសិកាម្តាយ (2n2c) ។
នៅក្នុងកោសិកាដែលមានជំងឺ ខូច ភាពចាស់ និងជាលិកាឯកទេសនៃរាងកាយ ដំណើរការបែងចែកខុសគ្នាបន្តិចអាចកើតឡើង - amitosis ។ អាមីតូស៊ីសហៅថាការបែងចែកដោយផ្ទាល់នៃកោសិកា eukaryotic ដែលការបង្កើតកោសិកាសមមូលហ្សែនមិនកើតឡើងទេ ដោយសារសមាសធាតុកោសិកាត្រូវបានចែកចាយមិនស្មើគ្នា។ វាកើតឡើងនៅក្នុងរុក្ខជាតិនៅក្នុង endosperm និងនៅក្នុងសត្វនៅក្នុងថ្លើម ឆ្អឹងខ្ចី និងជ្រុងនៃភ្នែក។
ម៉ីអូស៊ីស។ ដំណាក់កាលនៃជំងឺ meiosis
ម៉ីអូស៊ីស- នេះគឺជាវិធីសាស្រ្តនៃការបែងចែកដោយប្រយោលនៃកោសិកាដំណុះបឋម (2n2c) ដែលជាលទ្ធផលនៃកោសិកា haploid (1n1c) ដែលភាគច្រើនជាកោសិកាដំណុះត្រូវបានបង្កើតឡើង។
មិនដូច mitosis ទេ meiosis មានការបែងចែកកោសិកាបន្តបន្ទាប់គ្នាពីរដែលនីមួយៗនាំមុខដោយ interphase ។ ការបែងចែកដំបូងនៃ meiosis (meiosis I) ត្រូវបានគេហៅថា ការកាត់បន្ថយចាប់តាំងពីក្នុងករណីនេះចំនួនក្រូម៉ូសូមត្រូវបានកាត់បន្ថយពាក់កណ្តាលហើយការបែងចែកទីពីរ (meiosis II) - សមីការចាប់តាំងពីនៅក្នុងដំណើរការរបស់វា ចំនួននៃក្រូម៉ូសូមត្រូវបានអភិរក្ស។

អន្តរដំណាក់កាល Iដំណើរការស្រដៀងគ្នាទៅនឹង interphase នៃ mitosis ។ Meiosis Iចែកចេញជាបួនដំណាក់កាល៖ prophase I, metaphase I, anaphase I និង telophase I ។ ព្យាករណ៍ Iដំណើរការសំខាន់ពីរកើតឡើង៖ ការភ្ជាប់គ្នា និងការឆ្លងកាត់។ ការផ្សំ- នេះគឺជាដំណើរការនៃការលាយបញ្ចូលគ្នានៃក្រូម៉ូសូមដូចគ្នា (គូ) តាមបណ្តោយប្រវែងទាំងមូល។ គូនៃក្រូម៉ូសូមដែលបានបង្កើតឡើងកំឡុងពេលភ្ជាប់គ្នាត្រូវបានរក្សាទុករហូតដល់ចុងបញ្ចប់នៃ metaphase I ។
ឆ្លងកាត់- ការផ្លាស់ប្តូរគ្នាទៅវិញទៅមកនៃតំបន់ homologous នៃក្រូម៉ូសូម homologous ។ ជាលទ្ធផលនៃការឆ្លងកាត់ ក្រូម៉ូសូមដែលទទួលដោយសារពាង្គកាយពីឪពុកម្តាយទាំងពីរ ទទួលបាននូវការរួមផ្សំថ្មីនៃហ្សែន ដែលនាំទៅដល់ការលេចចេញនូវពូជពង្សចម្រុះហ្សែន។ នៅចុងបញ្ចប់នៃ prophase I ដូចជានៅក្នុង prophase នៃ mitosis នោះ nucleolus រលាយបាត់ centrioles បង្វែរទៅប៉ូលនៃកោសិកា ហើយស្រោមសំបុត្រនុយក្លេអ៊ែរក៏បែក។

IN metaphase Iគូនៃក្រូម៉ូសូមតម្រង់ជួរតាមបណ្តោយខ្សែអេក្វាទ័រនៃកោសិកា ដុំតូចៗនៃសរសៃពួរត្រូវបានភ្ជាប់ទៅកណ្តាលរបស់វា។
IN អាណាហ្វាស Iក្រូម៉ូសូម homologous ទាំងមូលមានក្រូម៉ាទីតពីរ បង្វែរទៅបង្គោល។
IN តេឡូហ្វាស Iនៅជុំវិញចង្កោមនៃក្រូម៉ូសូមនៅប៉ូលនៃកោសិកា ភ្នាសនុយក្លេអ៊ែរបង្កើតជាទម្រង់ nucleoli ។
Cytokinesis Iផ្តល់នូវការបែងចែក cytoplasms នៃកោសិកាកូនស្រី។
កោសិកាកូនស្រីដែលបង្កើតឡើងជាលទ្ធផលនៃ meiosis I (1n2c) មានលក្ខណៈតំណពូជដោយសារក្រូម៉ូសូមរបស់ពួកគេដែលបំបែកដោយចៃដន្យទៅកាន់ប៉ូលនៃកោសិកាមានហ្សែនមិនស្មើគ្នា។
លក្ខណៈប្រៀបធៀបនៃ mitosis និង meiosis
| សញ្ញា | មីតូស៊ីស | ម៉ីអូស៊ីស | |
| តើកោសិកាអ្វីខ្លះចាប់ផ្តើមបែងចែក? | Somatic (2n) | កោសិកាមេរោគបឋម (2n) | |
| ចំនួននៃការបែងចែក | 1 | 2 | |
| តើកោសិកាចំនួនប៉ុន្មាន និងប្រភេទណាខ្លះត្រូវបានបង្កើតឡើងក្នុងដំណើរការនៃការបែងចែក? | 2 somatic (2n) | 4 ផ្លូវភេទ (n) | |
| អន្តរដំណាក់កាល | ការរៀបចំកោសិកាសម្រាប់ការបែងចែក ការចម្លង DNA | ខ្លីណាស់ ការចម្លង DNA មិនកើតឡើងទេ។ | |
| ដំណាក់កាល | Meiosis I | ម៉ីអូស៊ីស II | |
| ប្រូហ្វាស | ការ condensation នៃក្រូម៉ូសូម ការបាត់ខ្លួននៃ nucleolus ការបែកបាក់នៃស្រោមសំបុត្រនុយក្លេអ៊ែរ ការភ្ជាប់ និងការឆ្លងកាត់អាចកើតឡើង | ការបង្រួបបង្រួមនៃក្រូម៉ូសូម ការបាត់ខ្លួននៃនុយក្លេអូល ការបែកបាក់នៃស្រោមសំបុត្រនុយក្លេអ៊ែរ | |
| metaphase | គូនៃក្រូម៉ូសូមមានទីតាំងនៅតាមខ្សែអេក្វាទ័រ ការបែងចែកត្រូវបានបង្កើតឡើង | ក្រូម៉ូសូមតម្រង់ជួរតាមខ្សែអេក្វាទ័រ ស្នូលនៃការបែងចែកត្រូវបានបង្កើតឡើង | |
| អាណាផាស | ក្រូម៉ូសូម homologous ពីក្រូម៉ាតពីរបង្វែរទៅបង្គោល | Chromatids បង្វែរទៅបង្គោល | |
| តេឡូហ្វាស | Chromosomes despiralize, ស្រោមសំបុត្រនុយក្លេអ៊ែរថ្មី និងទម្រង់ nucleoli | Chromosomes despiralize, ស្រោមសំបុត្រនុយក្លេអ៊ែរថ្មី និងទម្រង់ nucleoli | |
ដំណាក់កាលទី IIខ្លីណាស់ ចាប់តាំងពី DNA ទ្វេដងមិនកើតឡើងនៅក្នុងវា នោះគឺមិនមាន S-period ទេ។
ម៉ីអូស៊ីស IIក៏បែងចែកជាបួនដំណាក់កាល៖ ប្រូហ្វាសទី 2 មេតាហ្វាសទី 2 អាណាហ្វាសទី 2 និងតេឡូហ្វាសទី 2 ។ IN ដំណាក់កាលទី IIដំណើរការដូចគ្នាកើតឡើងដូចនៅក្នុងដំណាក់កាលទី 1 លើកលែងតែការភ្ជាប់គ្នា និងការឆ្លងកាត់។
IN metaphase IIក្រូម៉ូសូមមានទីតាំងនៅតាមខ្សែអេក្វាទ័រនៃកោសិកា។
IN អាណាហ្វាស IIក្រូម៉ូសូមបំបែកនៅកណ្តាល ហើយក្រូម៉ាទីតលាតសន្ធឹងឆ្ពោះទៅរកប៉ូល។
IN តេឡូហ្វាស IIភ្នាសនុយក្លេអ៊ែរ និង nucleoli បង្កើតនៅជុំវិញចង្កោមនៃក្រូម៉ូសូមកូនស្រី។
បន្ទាប់ពី cytokinesis IIរូបមន្តហ្សែននៃកោសិកាកូនស្រីទាំងបួនគឺ 1n1c ប៉ុន្តែពួកវាទាំងអស់មានសំណុំហ្សែនផ្សេងៗគ្នា ដែលជាលទ្ធផលនៃការឆ្លងកាត់ និងការបញ្ចូលគ្នាចៃដន្យនៃក្រូម៉ូសូមមាតា និងបិតានៅក្នុងកោសិកាកូនស្រី។
ការវិវត្តន៍នៃកោសិកាមេរោគនៅក្នុងរុក្ខជាតិ និងសត្វ
Gametogenesis(មកពីភាសាក្រិក។ gamete- ប្រពន្ធ gametes- ប្តីនិង ហ្សែន- ប្រភពដើម, ការកើតឡើង) គឺជាដំណើរការនៃការបង្កើតកោសិកាមេជីវិតឈ្មោល។
ដោយសារការបន្តពូជផ្លូវភេទភាគច្រើនទាមទារបុគ្គលពីរនាក់ - ស្ត្រី និងបុរស បង្កើតកោសិកាផ្លូវភេទខុសៗគ្នា - ស៊ុត និងមេជីវិតឈ្មោល បន្ទាប់មកដំណើរការនៃការបង្កើត gametes ទាំងនេះគួរតែខុសគ្នា។
ធម្មជាតិនៃដំណើរការនេះក៏អាស្រ័យទៅលើវិសាលភាពធំមួយផងដែរ ថាតើវាកើតឡើងនៅក្នុងកោសិការុក្ខជាតិ ឬសត្វទេ ព្រោះនៅក្នុងរុក្ខជាតិមានតែ mitosis ប៉ុណ្ណោះដែលកើតឡើងកំឡុងពេលបង្កើត gametes ខណៈពេលដែលនៅក្នុងសត្វទាំង mitosis និង meiosis កើតឡើង។
ការអភិវឌ្ឍនៃកោសិកាមេរោគនៅក្នុងរុក្ខជាតិ។នៅក្នុង angiosperms ការបង្កើតកោសិកាមេជីវិតឈ្មោលនិងស្ត្រីកើតឡើងនៅផ្នែកផ្សេងៗនៃផ្កា - stamens និង pistils រៀងគ្នា។
មុនពេលការបង្កើតកោសិកាមេជីវិតឈ្មោល - microgametogenesis(មកពីភាសាក្រិក។ មីក្រូ- តូច) - កើតឡើង microsporogenesisនោះគឺការបង្កើត microspores នៅក្នុង anthers នៃ stamens នេះ។ ដំណើរការនេះត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបែងចែក meiotic នៃកោសិកាម្តាយដែលបណ្តាលឱ្យមាន microspores haploid ចំនួនបួន។ Microgametogenesis ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបែងចែក mitotic នៃ microspore ដោយផ្តល់ឱ្យ gametophyte បុរសនៃកោសិកាពីរ - ធំមួយ។ បន្លែ(siphonogenic) និងរាក់ ជំនាន់. បន្ទាប់ពីការបែងចែក សត្វឈ្មោល gametophyte ត្រូវបានគ្របដណ្ដប់ដោយសំបកក្រាស់ និងបង្កើតជាគ្រាប់លំអង។ ក្នុងករណីខ្លះ សូម្បីតែនៅក្នុងដំណើរការនៃភាពចាស់ទុំនៃលំអង ហើយពេលខ្លះបានតែបន្ទាប់ពីការផ្ទេរទៅការមាក់ងាយនៃ pistil កោសិកាបង្កើតបានបែងចែកយ៉ាងសកម្មជាមួយនឹងការបង្កើតកោសិកាមេជីវិតឈ្មោលពីរដែលមិនមានចលនា - មេជីវិតឈ្មោល។. បន្ទាប់ពី pollination បំពង់ pollen មួយត្រូវបានបង្កើតឡើងពីកោសិកាលូតលាស់ ដែលតាមរយៈនោះ មេជីវិតឈ្មោលជ្រាបចូលទៅក្នុង ovary នៃ pistil សម្រាប់ការបង្កកំណើត។

ការបង្កើតកោសិកាមេជីវិតញីនៅក្នុងរុក្ខជាតិត្រូវបានគេហៅថា megagametogenesis(មកពីភាសាក្រិក។ មេហ្គា- ធំ) ។ វាកើតឡើងនៅក្នុង ovary នៃ pistil ដែលត្រូវបាននាំមុខដោយ megasporogenesisជាលទ្ធផលនៃការដែល megaspore បួនត្រូវបានបង្កើតឡើងពីកោសិកាមេនៃ megaspore ដែលស្ថិតនៅក្នុងស្នូលដោយការបែងចែក meiotic ។ មួយនៃ megaspores បែងចែក mitotically បីដងដែលផ្តល់ការកើនឡើងដល់ gametophyte ស្ត្រីដែលជាថង់អំប្រ៊ីយ៉ុងដែលមានស្នូលប្រាំបី។ ជាមួយនឹងភាពឯកោជាបន្តបន្ទាប់នៃ cytoplasms នៃកោសិកាកូនស្រី កោសិកាមួយក្នុងចំនោមកោសិកាលទ្ធផលបានក្លាយទៅជាស៊ុត ដែលនៅសងខាងដែលហៅថា synergids អង់ទីករបីត្រូវបានបង្កើតឡើងនៅចុងម្ខាងនៃថង់អំប្រ៊ីយ៉ុង ហើយនៅចំកណ្តាល។ ជាលទ្ធផលនៃការបញ្ចូលគ្នានៃស្នូល haploid ពីរកោសិកាកណ្តាល diploid ត្រូវបានបង្កើតឡើង។

ការវិវត្តនៃកោសិកាមេជីវិតនៅក្នុងសត្វ។នៅក្នុងសត្វ ដំណើរការពីរនៃការបង្កើតកោសិកាមេជីវិតឈ្មោលត្រូវបានសម្គាល់ - spermatogenesis និង oogenesis ។
ការបង្កើតមេជីវិតឈ្មោល។(មកពីភាសាក្រិក។ មេជីវិតឈ្មោល, មេជីវិតឈ្មោល។- គ្រាប់ពូជនិង ហ្សែន- ប្រភពដើម, ការកើតឡើង) គឺជាដំណើរការនៃការបង្កើតកោសិកាមេជីវិតឈ្មោលចាស់ទុំ - មេជីវិតឈ្មោល។ ចំពោះមនុស្ស វាកើតឡើងនៅក្នុងពងស្វាស ឬពងស្វាស ហើយចែកចេញជាបួនដំណាក់កាល៖ ការបន្តពូជ ការលូតលាស់ ភាពចាស់ទុំ និងការបង្កើត។

IN រដូវបង្កាត់ពូជកោសិកាមេជីវិតឈ្មោលចែកចេញជា mitotically ដែលជាលទ្ធផលនៅក្នុងការបង្កើត diploid spermatogonia. IN រយៈពេលលូតលាស់ spermatogonia ប្រមូលផ្តុំសារធាតុចិញ្ចឹមនៅក្នុង cytoplasm បង្កើនទំហំនិងប្រែទៅជា មេជីវិតឈ្មោលបឋម, ឬ spermatocytes នៃលំដាប់ទី 1. មានតែបន្ទាប់ពីនោះពួកគេចូលទៅក្នុង meiosis ( រយៈពេលទុំ) ដែលលទ្ធផលដំបូងជាពីរ មេជីវិតឈ្មោលបន្ទាប់បន្សំ, ឬ spermatocyte នៃលំដាប់ទី 2ហើយបន្ទាប់មក - កោសិកា haploid ចំនួនបួនដែលមានបរិមាណច្រើននៃ cytoplasm - មេជីវិតឈ្មោល។. IN រយៈពេលនៃការបង្កើតពួកវាបាត់បង់ស្ទើរតែទាំងអស់នៃ cytoplasm ហើយបង្កើតជា flagellum ប្រែទៅជាមេជីវិតឈ្មោល។
មេជីវិតឈ្មោល។, ឬ ស្ករកៅស៊ូ, - កោសិកាផ្លូវភេទបុរសចល័តតូចណាស់ដែលមានក្បាល ក និងកន្ទុយ។

IN ក្បាលលើកលែងតែស្នូលគឺ អាក្រូសូម- ស្មុគ្រស្មាញ Golgi ដែលបានកែប្រែ ដែលធានាការរំលាយភ្នាសនៃស៊ុតអំឡុងពេលបង្កកំណើត។ IN កមាន centrioles នៃមជ្ឈមណ្ឌលកោសិកា និងមូលដ្ឋាន កន្ទុយសេះបង្កើតជា microtubules ដែលគាំទ្រដោយផ្ទាល់នូវចលនារបស់មេជីវិតឈ្មោល។ វាក៏មានផ្ទុកសារធាតុ mitochondria ដែលផ្តល់ថាមពលដល់មេជីវិតឈ្មោល ATP សម្រាប់ចលនា។
Ovogenesis(មកពីភាសាក្រិក។ អ.ស.ប- ស៊ុតមួយនិង ហ្សែន- ប្រភពដើមការកើតឡើង) - នេះគឺជាដំណើរការនៃការបង្កើតកោសិកាមេជីវិតញីដែលចាស់ទុំ - ស៊ុត។ ចំពោះមនុស្ស វាកើតឡើងនៅក្នុងអូវែ ហើយមានបីដំណាក់កាល៖ ការបន្តពូជ ការលូតលាស់ និងការពេញវ័យ។ រយៈពេលនៃការបន្តពូជ និងការលូតលាស់ ស្រដៀងទៅនឹងការវិវត្តនៃ spermatogenesis កើតឡើងសូម្បីតែអំឡុងពេលមានការវិវឌ្ឍន៍នៃស្បូនក៏ដោយ។ ក្នុងពេលជាមួយគ្នានេះកោសិកា diploid ត្រូវបានបង្កើតឡើងពីកោសិកាដំណុះបឋមដែលជាលទ្ធផលនៃ mitosis ។ អូហ្គោនៀដែលបន្ទាប់មកប្រែទៅជា diploid បឋម oocytes, ឬ oocytes នៃលំដាប់ទី 1. Meiosis និង cytokinesis ជាបន្តបន្ទាប់ដែលកើតឡើងនៅក្នុង រយៈពេលទុំត្រូវបានកំណត់លក្ខណៈដោយការបែងចែកមិនស្មើគ្នានៃ cytoplasm នៃកោសិកាម្តាយ ដូច្នេះជាលទ្ធផលដំបូងគេទទួលបាន oocyte ទីពីរ, ឬ oocyte លំដាប់ទី 2, និង រាងប៉ូលដំបូងហើយបន្ទាប់មកពី oocyte ទីពីរ - ស៊ុតដែលរក្សាការផ្គត់ផ្គង់ទាំងមូលនៃសារធាតុចិញ្ចឹមនិងរាងកាយប៉ូលទីពីរខណៈពេលដែលរាងកាយប៉ូលដំបូងត្រូវបានបែងចែកជាពីរ។ សាកសពប៉ូល យកសម្ភារៈហ្សែនលើស។
នៅក្នុងមនុស្ស ស៊ុតត្រូវបានផលិតដោយមានចន្លោះពេល 28-29 ថ្ងៃ។ វដ្ដដែលជាប់ទាក់ទងនឹងភាពចាស់ទុំ និងការបញ្ចេញពងត្រូវបានគេហៅថា វដ្តរដូវ។
ស៊ុត- កោសិកាមេជីវិតញីដ៏ធំមួយ ដែលមិនត្រឹមតែផ្ទុកនូវសំណុំក្រូម៉ូសូម haploid ប៉ុណ្ណោះទេ ប៉ុន្តែថែមទាំងផ្គត់ផ្គង់សារធាតុចិញ្ចឹមសំខាន់ៗសម្រាប់ការអភិវឌ្ឍជាបន្តបន្ទាប់នៃអំប្រ៊ីយ៉ុង។

ស៊ុតនៅក្នុងថនិកសត្វត្រូវបានគ្របដណ្តប់ដោយភ្នាសចំនួន 4 ដែលកាត់បន្ថយលទ្ធភាពនៃការខូចខាតវាដោយកត្តាផ្សេងៗ។ អង្កត់ផ្ចិតនៃស៊ុតនៅក្នុងមនុស្សឈានដល់ 150-200 មីក្រូខណៈពេលដែលនៅក្នុងសត្វអូដ្ឋវាអាចមានច្រើនសង់ទីម៉ែត្រ។
ការបែងចែកកោសិកាគឺជាមូលដ្ឋានសម្រាប់ការលូតលាស់ ការអភិវឌ្ឍន៍ និងការបន្តពូជរបស់សារពាង្គកាយ។ តួនាទីរបស់ mitosis និង meiosis
ប្រសិនបើការបែងចែកកោសិកានៃសារពាង្គកាយ unicellular នាំឱ្យមានការកើនឡើងនៃចំនួនបុគ្គល ពោលគឺការបន្តពូជ បន្ទាប់មកនៅក្នុងសារពាង្គកាយពហុកោសិកា ដំណើរការនេះអាចមានអត្ថន័យខុសគ្នា។ ដូច្នេះ ការបែងចែកកោសិកានៃអំប្រ៊ីយ៉ុង ចាប់ផ្តើមពីហ្សីហ្គោត គឺជាមូលដ្ឋានជីវសាស្រ្តសម្រាប់ដំណើរការទំនាក់ទំនងគ្នានៃការលូតលាស់ និងការអភិវឌ្ឍន៍។ ការផ្លាស់ប្តូរស្រដៀងគ្នានេះត្រូវបានគេសង្កេតឃើញនៅក្នុងមនុស្សម្នាក់ក្នុងវ័យជំទង់នៅពេលដែលចំនួនកោសិកាមិនត្រឹមតែកើនឡើងប៉ុណ្ណោះទេប៉ុន្តែក៏មានការផ្លាស់ប្តូរគុណភាពនៅក្នុងខ្លួនផងដែរ។ ការបន្តពូជនៃសារពាង្គកាយពហុកោសិកាក៏ផ្អែកលើការបែងចែកកោសិកាផងដែរ ឧទាហរណ៍ ក្នុងអំឡុងពេលបន្តពូជដោយភេទ ដោយសារដំណើរការនេះ រាងកាយទាំងមូលត្រូវបានស្ដារឡើងវិញពីផ្នែកមួយនៃរាងកាយ ហើយក្នុងអំឡុងពេលបន្តពូជផ្លូវភេទ កោសិកាមេរោគត្រូវបានបង្កើតឡើងក្នុងអំឡុងពេល gametogenesis ដែលផ្តល់ជាបន្តបន្ទាប់។ សារពាង្គកាយថ្មី។ វាគួរតែត្រូវបានកត់សម្គាល់ថាវិធីសាស្រ្តសំខាន់នៃការបែងចែកកោសិកា eukaryotic - mitosis និង meiosis - មានសារៈសំខាន់ផ្សេងគ្នានៅក្នុងវដ្តជីវិតនៃសារពាង្គកាយ។
ជាលទ្ធផលនៃ mitosis មានការចែកចាយឯកសណ្ឋាននៃសម្ភារៈតំណពូជរវាងកោសិកាកូនស្រី - ច្បាប់ចម្លងពិតប្រាកដរបស់ម្តាយ។ បើគ្មាន mitosis ទេ អត្ថិភាព និងការលូតលាស់នៃសារពាង្គកាយពហុកោសិកាដែលវិវឌ្ឍន៍ចេញពីកោសិកាតែមួយ ហ្សីហ្គោត នឹងមិនអាចទៅរួចទេ ព្រោះកោសិកាទាំងអស់នៃសារពាង្គកាយបែបនេះត្រូវតែមានព័ត៌មានហ្សែនដូចគ្នា។
នៅក្នុងដំណើរការនៃការបែងចែកកោសិកាកូនស្រីកាន់តែមានភាពចម្រុះនៅក្នុងរចនាសម្ព័ន្ធ និងមុខងារ ដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការធ្វើឱ្យសកម្មនៃក្រុមថ្មីនៃហ្សែននៅក្នុងពួកវាដោយសារតែអន្តរកម្មរវាងកោសិកា។ ដូច្នេះ mitosis គឺចាំបាច់សម្រាប់ការអភិវឌ្ឍនៃសារពាង្គកាយមួយ។
វិធីសាស្រ្តនៃការបែងចែកកោសិកានេះគឺចាំបាច់សម្រាប់ដំណើរការនៃការបន្តពូជ និងការបង្កើតឡើងវិញ (ការស្តារឡើងវិញ) នៃជាលិកាដែលខូច ក៏ដូចជាសរីរាង្គផងដែរ។
Meiosis ជាវេនធានានូវភាពស្ថិតស្ថេរនៃ karyotype អំឡុងពេលបន្តពូជផ្លូវភេទ ព្រោះវាកាត់បន្ថយពាក់កណ្តាលនៃក្រូម៉ូសូមមុនពេលបន្តពូជផ្លូវភេទ ដែលបន្ទាប់មកត្រូវបានស្តារឡើងវិញជាលទ្ធផលនៃការបង្កកំណើត។ លើសពីនេះទៀត meiosis នាំទៅរកការលេចឡើងនៃការរួមបញ្ចូលគ្នាថ្មីនៃហ្សែនឪពុកម្តាយដោយសារតែការឆ្លងកាត់និងការបញ្ចូលគ្នាចៃដន្យនៃក្រូម៉ូសូមនៅក្នុងកោសិកាកូនស្រី។ អរគុណចំពោះបញ្ហានេះ ពូជពង្សមានភាពចម្រុះហ្សែន ដែលផ្តល់សម្ភារៈសម្រាប់ការជ្រើសរើសធម្មជាតិ និងជាមូលដ្ឋានសម្ភារៈនៃការវិវត្តន៍។ ការផ្លាស់ប្តូរចំនួន រូបរាង និងទំហំនៃក្រូម៉ូសូមនៅលើដៃម្ខាងអាចនាំឱ្យមានរូបរាងនៃគម្លាតផ្សេងៗក្នុងការវិវត្តនៃសារពាង្គកាយ និងសូម្បីតែការស្លាប់របស់វា ហើយម្យ៉ាងវិញទៀត វាអាចនាំឱ្យមានរូបរាងរបស់មនុស្សម្នាក់ៗ។ កាន់តែសម្របខ្លួនទៅនឹងបរិស្ថាន។
ដូច្នេះ កោសិកាគឺជាឯកតានៃការលូតលាស់ ការអភិវឌ្ឍន៍ និងការបន្តពូជរបស់សារពាង្គកាយ។
សំណួរទី 1។
កោសិកានេះមានធាតុគីមីប្រហែល 80 នៃប្រព័ន្ធតាមកាលកំណត់របស់ D. I. Mendeleev ។ ធាតុទាំងអស់នេះក៏ត្រូវបានរកឃើញផងដែរនៅក្នុងធម្មជាតិគ្មានជីវិត ដែលដើរតួជាភស្តុតាងមួយនៃភាពសាមញ្ញនៃធម្មជាតិដែលមានចលនា និងគ្មានជីវិត។ ទោះជាយ៉ាងណាក៏ដោយ សមាមាត្រនៃធាតុគីមីនៅក្នុងសារពាង្គកាយមានជីវិតគឺខុសពីវត្ថុនៃធម្មជាតិគ្មានជីវិត។ នៅក្នុងសារពាង្គកាយមានជីវិត ធាតុភាគច្រើនស្ថិតក្នុងទម្រង់ជាសមាសធាតុគីមី សារធាតុរំលាយក្នុងទឹក មានតែសារពាង្គកាយមានជីវិតប៉ុណ្ណោះដែលមានសារធាតុសរីរាង្គ៖ ប្រូតេអ៊ីន ខ្លាញ់ កាបូអ៊ីដ្រាត និងអាស៊ីត nucleic
សំណួរទី 2 ។
សមាសធាតុគីមីកោសិការុក្ខជាតិនិងសត្វគឺស្រដៀងគ្នា។ សារពាង្គកាយមានជីវិតទាំងអស់ត្រូវបានផ្សំឡើងដោយធាតុដូចគ្នា សមាសធាតុអសរីរាង្គ និងសរីរាង្គ។ ប៉ុន្តែមាតិកានៃធាតុផ្សេងគ្នានៅក្នុងកោសិកាផ្សេងគ្នាគឺខុសគ្នា។ ប្រភេទកោសិកានីមួយៗមានលេខផ្សេងគ្នានៃម៉ូលេគុលសរីរាង្គជាក់លាក់។ នៅក្នុងកោសិការុក្ខជាតិ កាបូអ៊ីដ្រាតស្មុគ្រស្មាញ (ជាតិសរសៃ ម្សៅ) គ្របដណ្ដប់ ហើយនៅក្នុងសត្វមានប្រូតេអ៊ីន និងខ្លាញ់ច្រើន។ ក្រុមនីមួយៗនៃសារធាតុសរីរាង្គ (ប្រូតេអ៊ីន កាបូអ៊ីដ្រាត ខ្លាញ់ អាស៊ីតនុយក្លេអ៊ីក) នៅក្នុងប្រភេទកោសិកាណាមួយអនុវត្តមុខងាររបស់វាផ្ទាល់ (ការផ្ទុកអាស៊ីតនុយក្លេអ៊ីក និងការបញ្ជូនព័ត៌មានតំណពូជ កាបូអ៊ីដ្រាត - ថាមពល។ល។)។
សំណួរទី 3 ។
ធាតុជាច្រើននៃប្រព័ន្ធតាមកាលកំណត់របស់ Mendeleev ត្រូវបានរកឃើញនៅក្នុងកោសិកា។ មុខងារចំនួន ២៧ ត្រូវបានកំណត់។ ធម្មតាបំផុតគឺកាបូន អ៊ីដ្រូសែន អាសូត អុកស៊ីហ្សែន ផូស្វ័រ និងស្ពាន់ធ័រ។ ពួកវាបង្កើតបាន 99% នៃម៉ាសកោសិកាសរុប។
ធាតុគីមីដែលបង្កើតកោសិកា ចែកចេញជាបីក្រុម៖ macronutrients, ធាតុដាន, មីក្រូរ៉ែ.
1. ម៉ាក្រូសារធាតុចិញ្ចឹម: C, H, N, Ca, K, Mg, Na, Fe, S, P, C1 ។ ធាតុទាំងនេះមានច្រើនជាង 99% នៃម៉ាសកោសិកាសរុប។ ពួកគេខ្លះផ្តោតអារម្មណ៍ខ្លាំង។ បរិមាណអុកស៊ីសែន 65-75%; កាបូន - 15-18%; អាសូត - 1.5-3% ។
2. ធាតុដាន៖ Cu, B, Co, Mo, Mn, Ni, Br, I និងអ្នកដទៃ។ ចំណែករបស់ពួកគេនៅក្នុងក្រឡាក្នុងគណនីសរុបមានច្រើនជាង 0.1%; ការប្រមូលផ្តុំនីមួយៗមិនលើសពី 0,001% ។ ទាំងនេះគឺជាអ៊ីយ៉ុងដែកដែលជាផ្នែកមួយនៃសារធាតុសកម្មជីវសាស្រ្ត (អ័រម៉ូន អង់ស៊ីម ជាដើម)។ ឧទាហរណ៍ cobalt គឺជាផ្នែកមួយនៃវីតាមីន BO, C, H, N, Ca, K, Mg, Na, Fe 12 ដែលពាក់ព័ន្ធនឹង hematopoiesis ហើយ fluorine ស្ថិតនៅក្នុងកោសិកានៃស្រទាប់ធ្មេញ។
3. ធាតុអ៊ុលត្រាសោ: អ៊ុយរ៉ាញ៉ូម, មាស, បេរីលញ៉ូម, បារត, សេសេញ៉ូម, សេលេញ៉ូមនិងផ្សេងទៀត។ ការផ្តោតអារម្មណ៍របស់ពួកគេមិនលើសពី 0.000001% ។ តួនាទីសរីរវិទ្យានៃពួកគេជាច្រើនមិនត្រូវបានបង្កើតឡើងទេ។
សំណួរទី 4 ។
សមាសធាតុសរីរាង្គបង្កើតបានជាមធ្យម 10% នៃម៉ាស់កោសិកានៃសារពាង្គកាយមានជីវិត។ ទាំងនេះរួមមានប៉ូលីម៊ែរជីវសាស្រ្ត - ប្រូតេអ៊ីន អាស៊ីត nucleic និងកាបូអ៊ីដ្រាត ក៏ដូចជាខ្លាញ់ និងម៉ូលេគុលតូចៗមួយចំនួន -
សំណួរទី 5 ។
កំប្រុក- សារធាតុសរីរាង្គវត្ថុធាតុ polymeric ម៉ូលេគុលខ្ពស់ ដែលកំណត់រចនាសម្ព័ន្ធ និងសកម្មភាពសំខាន់នៃកោសិកា និងសារពាង្គកាយទាំងមូល។ ប្រូតេអ៊ីនបង្កើតបាន 10-18% នៃម៉ាសកោសិកាសរុប។
ប្រូតេអ៊ីនអនុវត្តមុខងារដូចខាងក្រោមៈ
អង់ស៊ីម (ឧទាហរណ៍អាមីឡាសបំបែកកាបូអ៊ីដ្រាត);
រចនាសម្ព័ន្ធ (ឧទាហរណ៍ពួកវាជាផ្នែកមួយនៃភ្នាសកោសិកា);
អ្នកទទួល (ឧទាហរណ៍ rhodopsin លើកកម្ពស់ចក្ខុវិស័យកាន់តែប្រសើរ);
ការដឹកជញ្ជូន (ឧទាហរណ៍អេម៉ូក្លូប៊ីនដឹកអុកស៊ីសែនឬកាបូនឌីអុកស៊ីត);
ការពារ (ឧទាហរណ៍ immunoglobulins ត្រូវបានចូលរួមនៅក្នុងការបង្កើតភាពស៊ាំ);
ម៉ូទ័រ (ឧទាហរណ៍ actin, myosin, ត្រូវបានចូលរួមនៅក្នុងការកន្ត្រាក់នៃសរសៃសាច់ដុំ);
អរម៉ូន (ឧទាហរណ៍អាំងស៊ុយលីនបំលែងគ្លុយកូសទៅជា glycogen);
ថាមពល (នៅពេលបំបែកប្រូតេអ៊ីន 1 ក្រាមថាមពល 4.2 kcal ត្រូវបានបញ្ចេញ) ។
សំណួរទី 6 ។
កាបូអ៊ីដ្រាតដើរតួជាប្រភពថាមពលសំខាន់នៅក្នុងកោសិកា។ នៅក្នុងដំណើរការនៃការកត់សុីនៃកាបូអ៊ីដ្រាត 1 ក្រាមថាមពល 17,6 kJ ត្រូវបានបញ្ចេញ។ ម្សៅនៅក្នុងរុក្ខជាតិ និង glycogen នៅក្នុងសត្វ ដែលត្រូវបានដាក់ក្នុងកោសិកា បម្រើជាទុនបម្រុងថាមពល។ សារពាង្គកាយមានជីវិតអាចផ្ទុកកាបូអ៊ីដ្រាតក្នុងទម្រង់ជាម្សៅ (នៅក្នុងរុក្ខជាតិ) និង glycogen (នៅក្នុងសត្វ និងផ្សិត)។ នៅក្នុងមើមដំឡូង ម្សៅអាចបង្កើតបានរហូតដល់ 80% នៃម៉ាស ហើយសត្វមានកាបូអ៊ីដ្រាតច្រើន ជាពិសេសនៅក្នុងកោសិកាថ្លើម និងសាច់ដុំ រហូតដល់ 5%។
កាបូអ៊ីដ្រាតក៏អនុវត្តមុខងារផ្សេងទៀតផងដែរដូចជាការគាំទ្រនិងការការពារ។ ជាឧទាហរណ៍ សែលុយឡូសបង្កើតជាជញ្ជាំងនៃកោសិការុក្ខជាតិ៖ ប៉ូលីស្យូមស្មុគ្រស្មាញ ឈីទីន- សមាសធាតុរចនាសម្ព័ន្ធសំខាន់នៃគ្រោងខាងក្រៅនៃ arthropods ។ Chitin ក៏អនុវត្តមុខងារសាងសង់នៅក្នុងផ្សិតផងដែរ។ ពួកវាមានវត្តមាននៅក្នុង DNA, RNA និង ATP ក្នុងទម្រង់ជា deoxyribose និង ribose។
សំណួរទី 7 ។
ខ្លាញ់អនុវត្តមុខងារមួយចំនួននៅក្នុងរាងកាយ៖
រចនាសម្ព័ន្ធ (ចូលរួមក្នុងការសាងសង់ភ្នាស);
ថាមពល (នៅពេលដែលរាងកាយបំបែកជាតិខ្លាញ់ 1 ក្រាម 9,2 kcal នៃថាមពលត្រូវបានបញ្ចេញ - 2,5 ដងច្រើនជាងពេលដែលបរិមាណកាបូអ៊ីដ្រាតដូចគ្នាបំបែក);
ការពារ (ពីការបាត់បង់កំដៅ, ការខូចខាតមេកានិច);
ខ្លាញ់គឺជាប្រភពនៃទឹក endogenous (នៅពេលដែល 10 ក្រាមនៃជាតិខ្លាញ់ត្រូវបានកត់សុី, 11 ក្រាមនៃទឹកត្រូវបានបញ្ចេញ) ។ នេះគឺមានសារៈសំខាន់ខ្លាំងណាស់សម្រាប់ការ hibernating សត្វ - កំប្រុកដី, marmots: ដោយសារតែទុនបម្រុងជាតិខ្លាញ់ subcutaneous របស់ពួកគេពួកគេមិនអាចផឹកនៅពេលនេះរហូតដល់ពីរខែ។ ក្នុងអំឡុងពេលឆ្លងកាត់វាលខ្សាច់សត្វអូដ្ឋទៅដោយមិនផឹករហូតដល់ពីរសប្តាហ៍ - ពួកគេបានទាញយកទឹកដែលចាំបាច់សម្រាប់រាងកាយពីខ្ទមរបស់ពួកគេ - បង្កាន់ដៃនៃជាតិខ្លាញ់។
បទប្បញ្ញត្តិនៃការរំលាយអាហារ (ឧទាហរណ៍អរម៉ូនស្តេរ៉ូអ៊ីត - corticosterone ជាដើម) ។
សំណួរទី 8 ។
សមាសធាតុអសរីរាង្គទូទៅបំផុតនៅក្នុងសារពាង្គកាយមានជីវិតគឺទឹក។ មាតិការបស់វានៅក្នុងកោសិកានៃប្រភេទផ្សេងៗគ្នាមានភាពខុសប្លែកគ្នាយ៉ាងទូលំទូលាយ: នៅក្នុងកោសិកានៃ enamel ធ្មេញទឹកគឺប្រហែល 10% ហើយនៅក្នុងកោសិកានៃអំប្រ៊ីយ៉ុងគុណ - ច្រើនជាង 90% ។ នៅក្នុងខ្លួនរបស់ចាហួយ ទឹកមានដល់ទៅ ៩៨%។ ប៉ុន្តែជាមធ្យម ក្នុងសារពាង្គកាយពហុកោសិកា ទឹកបង្កើតបានប្រហែល 80% នៃទំងន់រាងកាយ។ មុខងារចម្បងរបស់វាមានដូចខាងក្រោម៖
1. សារធាតុរំលាយជាសកល។
2. បរិយាកាសដែលប្រតិកម្មជីវគីមីកើតឡើង។
3. កំណត់លក្ខណៈសរីរវិទ្យានៃកោសិកា (ភាពបត់បែន បរិមាណរបស់វា)។
4. ចូលរួមក្នុងប្រតិកម្មគីមី។
5. រក្សាតុល្យភាពកម្ដៅនៃកោសិកា និងសារពាង្គកាយទាំងមូល ដោយសារសមត្ថភាពកំដៅខ្ពស់ និងចរន្តកំដៅ។
6. មធ្យោបាយសំខាន់សម្រាប់ការដឹកជញ្ជូនសារធាតុ។
សំណួរទី 9 ។
កាបូអ៊ីដ្រាតរួមមានសមាសធាតុសរីរាង្គធម្មជាតិដូចខាងក្រោមៈ គ្លុយកូស ហ្វ្រូតូស ស៊ូក្រូស ម៉ុលតូស ឡាក់តូស រីបូស ដេអុកស៊ីរីបូស ឈីទីន ម្សៅ គ្លីកូហ្សែន និងសែលុយឡូស។
សំណួរទី 10 ។
តម្លៃនៃអាស៊ីត nucleic គឺខ្ពស់ណាស់។ លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធគីមីរបស់ពួកគេផ្តល់នូវលទ្ធភាពនៃការរក្សាទុកផ្ទេរទៅស៊ីតូប្លាសនិងទទួលមរតកទៅកោសិកាកូនស្រីព័ត៌មានអំពីរចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅក្នុងកោសិកានីមួយៗ។ ពួកវាជាផ្នែកមួយនៃក្រូម៉ូសូម - រចនាសម្ព័ន្ធពិសេសដែលមានទីតាំងនៅស្នូលកោសិកា។ អាស៊ីតនុយក្លេអ៊ីកក៏ត្រូវបានរកឃើញផងដែរនៅក្នុង cytoplasm និងសរីរាង្គរបស់វា។
សំណួរទី 11 ។
ស៊ីលីកុន អាលុយមីញ៉ូម អុកស៊ីហ្សែន និងសូដ្យូម គឺជារឿងធម្មតាបំផុតនៅក្នុងសំបកផែនដី (ប្រហែល 90%) ។ នៅក្នុងសារពាង្គកាយមានជីវិតប្រហែល 98% នៃម៉ាស់ត្រូវបានបង្កើតឡើងដោយធាតុបួនគឺ អ៊ីដ្រូសែន អុកស៊ីសែន កាបូន និងអាសូត។ ភាពខុសគ្នានេះគឺដោយសារតែភាពបារម្ភនៃលក្ខណៈសម្បត្តិគីមីនៃធាតុដែលបានរាយបញ្ជីដែលជាលទ្ធផលដែលពួកគេបានប្រែទៅជាសមរម្យបំផុតសម្រាប់ការបង្កើតម៉ូលេគុលដែលបំពេញមុខងារជីវសាស្រ្ត។ អ៊ីដ្រូសែន អុកស៊ីហ្សែន កាបូន និងអាសូត អាចបង្កើតចំណងគីមីដ៏រឹងមាំ ដែលបណ្តាលឱ្យមានសមាសធាតុគីមីជាច្រើនប្រភេទ។ សមាសភាពនៃសារពាង្គកាយមានជីវិតរួមមានសារធាតុសរីរាង្គ (ប្រូតេអ៊ីន ខ្លាញ់ កាបូអ៊ីដ្រាត អាស៊ីតនុយក្លេអ៊ីក) និងសារធាតុអសរីរាង្គ (ទឹក អំបិលរ៉ែ)។







