I evolutionsprocessen sker (resultaten av evolutionen):
- Förändring, komplikation av organismer.
- Uppkomsten av nya arter(ökning av antalet [mångfald] arter).
- Anpassning av organismer till miljöförhållanden (till levnadsförhållanden), till exempel:
- skadedjursresistens mot bekämpningsmedel,
- ökenväxters motståndskraft mot torka,
- växternas anpassningsförmåga till insektspollinering,
- varning (ljus) färg hos giftiga djur,
- mimik (imitation av ett ofarligt djur till ett farligt),
- skyddande färg och form (osynlighet i bakgrunden).
All kondition är relativ, dvs. anpassar kroppen till endast en vissa villkor... När förutsättningarna förändras kan konditionen bli värdelös eller till och med skadlig (en mörk mal på en miljövänlig björk).
Befolkning är en enhet av evolution
En population är en samling individer av samma art, som lever under lång tid i en viss del av området (elementär strukturell formens enhet).
Inom befolkningen fri korsning, är korsningen mellan populationer begränsad.
Populationer av en art skiljer sig något från varandra, eftersom naturligt urval anpassar varje population till de specifika förhållandena i dess utbredningsområde (population - evolutionens enhet).
Mikroevolution och makroevolution
Mikroevolution- dessa är förändringar som sker i populationer under påverkan av evolutionens drivkrafter. I slutändan leder till uppkomsten av en ny art.
Makrovolution- Detta är processen för bildandet av stora systematiska enheter, supraspecifika taxa - släkten, familjer och högre.
Upprätta en överensstämmelse mellan egenskaperna hos det biologiska systemet och det system för vilket dessa egenskaper är karakteristiska: 1) population, 2) arter. Skriv ner siffrorna 1 och 2 i rätt ordning.
A) en elementär utvecklingsenhet i enlighet med STE
B) representanter får aldrig träffas på grund av isolering
C) Sannolikheten för korsning mellan medlemmar i gruppen är så sannolik som möjligt
D) delas upp i mindre, separata grupper
E) utbredningsområdet kan täcka flera kontinenter
Svar
Välj den som är mest korrekt. Framväxten av anpassning av arter till miljön är resultatet av
1) utseendet på ändringar ändras
2) interaktion mellan evolutionära faktorer
3) komplicera deras organisation
4) biologiska framsteg
Svar
Svar
Upprätta en överensstämmelse mellan evolutionens egenskaper och dess särdrag: 1) faktor, 2) resultat
A) naturligt urval
B) organismernas anpassningsförmåga till miljön
C) bildandet av nya arter
D) kombinativ variabilitet
E) bevarande av arter under stabila förhållanden
E) kampen för tillvaron
Svar
Välj den som är mest korrekt. Resultatet av växternas makroevolution är uppkomsten av nya
1) typer
2) avdelningar
3) befolkningar
4) sorter
Svar
Välj den som är mest korrekt. En elementär evolutionär enhet betraktas
1) visa
2) genom
3) befolkning
4) genotyp
Svar
Välj den som är mest korrekt. Historisk förändring kallas makroevolution
1) biocenoser
2) befolkningar
3) supraspecifika taxa
4) typer
Svar
Upprätta en överensstämmelse mellan egenskaperna hos den evolutionära processen och den nivå av evolution på vilken den sker: 1) mikroevolutionär, 2) makroevolutionär. Skriv ner siffrorna 1 och 2 i rätt ordning.
A) nya arter bildas
B) supraspecifika taxa bildas
C) populationens genpool förändras
D) framsteg uppnås genom idioadaptation
E) framsteg uppnås genom aromorfoser eller degeneration
Svar
Läs texten. Välj tre sanna påståenden. Skriv ner siffrorna under vilka de anges.(1) Den evolutionära processen består av makroevolution och mikroevolution. (2) Mikroevolution sker på populations-artnivå. (3) Evolutionens styrande faktor är kampen för tillvaron. (4) Evolutionens elementära enhet är en klass. (5) De huvudsakliga formerna av naturligt urval är drivande, stabiliserande, störande.
Svar
Välj den som är mest korrekt. Konsekvensen av organismers utveckling kan inte övervägas
1) organismers anpassningsförmåga till miljön
2) mångfalden i den organiska världen
3) ärftlig variation
4) bildandet av nya arter
Svar
Välj den som är mest korrekt. Evolutionsresultat inkluderar
1) organismernas variation
2) ärftlighet
3) anpassningsförmåga till miljöförhållanden
4) naturligt urval av ärftliga förändringar
Svar
Välj den som är mest korrekt. Det skiktade arrangemanget av trädens rotsystem i skogen är en anpassning som bildades under påverkan av
1) metabolism
2) cirkulation av ämnen
3) evolutionens drivkrafter
4) självreglering
Svar
Välj tre teser av den syntetiska evolutionsteorin.
1) evolutionens enhet - befolkning
2) evolutionens enhet - art
3) evolutionsfaktorer - mutationsvariabilitet, gendrift, befolkningsvågor
4) evolutionens faktorer - ärftlighet, föränderlighet, kamp för tillvaron
5) former av naturligt urval - motiv och sexuellt
6) former av naturligt urval - drivande, stabiliserande, störande
Svar
Läs texten. Välj tre meningar som beskriver exempel på mimik i naturen. Skriv ner siffrorna under vilka de anges. (1) Fågelhonor som häckar på marken smälter praktiskt taget samman med områdets allmänna bakgrund. (2) Ägg och kycklingar är också osynliga hos dessa fågelarter. (3) Många icke-giftiga ormar är mycket lika giftormar. (4) Ett antal stickande eller insekter med giftkörtlar utvecklar ljusa färger, vilket avskräcker någon från att prova dem. (5) Bin och deras imitatorer - svävflugor - är oattraktiva för insektsätande fåglar. (6) Hos vissa fjärilar liknar mönstret på vingarna ögonen på ett rovdjur.
Svar
Läs texten. Välj tre meningar som beskriver exempel på skyddande färgning i naturen. Skriv ner siffrorna under vilka de anges. (1) Honor som häckar på marken smälter praktiskt taget samman med områdets allmänna bakgrund, och deras ägg och ungar är också osynliga. (2) Likheten med miljöobjekt tillåter också många djur att undvika möten med rovdjur. (3) Ett antal stickande eller giftiga insekter utvecklar ljusa färger som avskräcker någon från att prova dem. (4) I regionerna i Fjärran Norden är vit färg mycket vanlig bland djur. (5) Hos vissa fjärilar liknar mönstret på vingarna ögonen på ett rovdjur. (6) Hos vissa djur efterliknar fläckig färg växlingen av ljus och skugga i den omgivande naturen och gör dem mindre synliga i täta snår.
Svar
Betrakta teckningen av en björkfjäril och bestäm (A) typen av anpassning, (B) formen av naturligt urval och (C) riktningen för utvecklingen som ledde till bildandet av två former av fjärilar. Skriv ner de tre siffrorna (termnummer från listan) i rätt ordning.
1) idioanpassning
2) mimik
3) konvergens
4) körning
5) aromorfos
6) förklädnad
7) stabilisering
Svar

Betrakta en ritning som visar frambenen på olika däggdjur och bestäm (A) evolutionens riktning, (B) mekanismen för evolutionär transformation, (C) formen av naturligt urval som ledde till bildandet av sådana organ. Välj lämplig term från listan för varje bokstav.
1) aromorfos
2) stabilisering
3) allmän degeneration
4) divergens
5) körning
6) idioanpassning
7) morfofysiologisk regression
8) konvergens
Svar

Tänk på ritningen av en sjöhäst och bestäm (A) typen av anpassning, (B) formen av naturligt urval och (C) den evolutionära vägen som ledde till bildandet av en sådan anpassning hos detta djur. Välj lämplig term från listan för varje bokstav.
1) spricker
2) förklädnad
3) idioanpassning
4) styckad färgning
5) parallellitet
6) mimetism
7) körning
Svar

Betrakta teckningen av en kolibri och en myrslok och bestäm (A) typen av anpassning, (B) formen av naturligt urval och (C) den evolutionära vägen som ledde till bildandet av sådana anpassningar. Välj lämplig term från listan för varje bokstav.
1) körning
2) störande
3) degeneration
4) idioanpassning
5) divergens
6) specialisering
7) sexuell dimorfism
Svar
Välj två rätta svar av fem och skriv ner siffrorna under vilka de anges. Vilka processer äger rum på befolkningsnivå?
1) ontogenes
2) divergens
3) embryogenes
4) aromorfos
5) fri överfart
Svar
Läs texten. Välj tre meningar som beskriver befolkningen som en enhet för utveckling. Skriv ner siffrorna under vilka de anges. (1) En art är en samling av populationer. (2) En populations huvudsakliga egenskaper är genetisk mångfald och förändring över tid. (3) Populationer av en art skiljer sig åt i antal, täthet, ålder och könsstruktur. (4) Varje population upptar en del av arternas utbredningsområde. (5) Det finns en konstant mutationsprocess i befolkningen, och den fördelaktiga mutationen sprider sig. (6) Inom en population utbyts gener mellan individer som ett resultat av fri korsning.
Svar
Läs texten. Välj tre meningar som korrekt karakteriserar befolkningen som en evolutionsenhet i den organiska världen. Skriv ner siffrorna under vilka de anges. (1) En population är en samling fritt korsande individer som har bott i ett gemensamt territorium under lång tid. (2) Populationens huvudsakliga egenskaper är storlek, täthet, ålder, kön, rumslig struktur, vilket gör att individer fritt kan korsa sig och ge fertil avkomma. (3) Befolkningen är en strukturell enhet i biosfären. (4) En befolkning är en elementär enhet i den organiska världens taxonomi. (5) Larverna från olika insekter som lever i en sötvattenförekomst utgör en population. (6) I befolkningen dör vissa individer och andra överlever, därför existerar den under lång tid.
Svar
Välj tre alternativ. Vilket av följande exempel illustrerar resultaten av utvecklingen av den organiska världen?
1) skyddande färg på isbjörnen, ripa som lever i norr
2) kampen för överlevnad mellan tallar och granar i skogen
3) mänsklig förädling av nya sorter av växter och djurraser
4) varning (hotande) färgning av nyckelpigor
5) stoppa flödet av gener från population till population (isolering)
6) likheten mellan kroppsform och färg hos oskyddade djur med skyddade
Svar

Analysera tabellen "Fitness of organisms". Välj lämplig term från listan för varje bokstav.
1) växling av ljusa fläckar, ränder, kroppsdelar
2) smälter samman med miljöns huvudbakgrund
3) döljer objektet mot bakgrund av streck av ljus och skugga
4) mimik
5) knutlik eller bladformad
6) nyckelpiga skalbagge, firebug, pilgiftsgrodor
7) glasfjäril, svävfluga
8) stick insekt, bönsyrsa
Svar
© D.V. Pozdnyakov, 2009-2019
Snäll, hans kriterier. En population är en strukturell enhet av en art och en elementär enhet av evolution. Mikroevolution. Bildande av nya arter. Specifikationsmetoder. Bevarande av arternas mångfald som grund för biosfärens hållbarhet
Visa, dess kriterier
Grundaren av den moderna taxonomin K. Linnaeus ansåg arten som en grupp organismer liknande morfologiska egenskaper, som fritt korsar sig med varandra. Med utvecklingen av biologin erhölls bevis för att skillnaderna mellan arter är mycket djupare och påverkar den kemiska sammansättningen och koncentrationen av ämnen i vävnader, riktningen och hastigheten för kemiska reaktioner, naturen och intensiteten av vitala processer, antalet och formen av kromosomer, det vill säga arten är den minsta gruppen av organismer som återspeglar deras nära släktskap. Dessutom existerar inte arter för alltid – de uppstår, utvecklas, ger upphov till nya arter och försvinner.
Seär en uppsättning individer som liknar strukturen och egenskaperna hos vitala processer, som har ett gemensamt ursprung, fritt förökar sig med varandra i naturen och ger fertil avkomma.
Alla individer av samma art har samma karyotyp och upptar ett visst geografiskt område i naturen - område.
Tecken på likhet mellan individer av samma art kallas formulärets kriterier... Eftersom inget av kriterierna är absolut måste en uppsättning kriterier användas för att korrekt bestämma arten.
De viktigaste artkriterierna är morfologiska, fysiologiska, biokemiska, ekologiska, geografiska, etologiska (beteendemässiga) och genetiska.
- Morfologiska- en uppsättning externa och inre tecken organismer av samma art. Medan vissa arter har unika egenskaper, är morfologiska egenskaper ofta mycket svåra att skilja mellan närbesläktade arter. Så nyligen har ett antal syskonarter upptäckts som lever på samma territorium, till exempel hus- och barrmöss, därför är det oacceptabelt att enbart använda det morfologiska kriteriet för att bestämma arten.
- Fysiologisk- likheten mellan processerna för vital aktivitet i organismer, först och främst reproduktion. Det är inte heller universellt, eftersom vissa arter i naturen korsar sig och ger fertil avkomma.
- Biokemisk- likheten mellan den kemiska sammansättningen och förloppet av metaboliska processer. Trots det faktum att dessa indikatorer kan variera avsevärt hos olika individer av samma art, ägnas för närvarande stor uppmärksamhet åt dem, eftersom egenskaperna hos strukturen och sammansättningen av biopolymerer hjälper till att identifiera arter även på molekylär nivå och fastställa graden av deras relation.
- Ekologisk- skillnaden i arter beroende på deras tillhörighet till vissa ekosystem och ekologiska nischer som de upptar. Men många obesläktade arter upptar liknande ekologiska nischer, så detta kriterium kan användas för att särskilja en art endast i kombination med andra karaktärer.
- Geografisk- förekomsten av en population av varje art i en viss del av biosfären - ett område som skiljer sig från områdena för alla andra arter. På grund av det faktum att för många arter gränserna för utbredningsområdena sammanfaller, och det finns också ett antal kosmopolitiska arter, vars utbredning täcker stora områden, kan det geografiska kriteriet inte heller fungera som ett "specifikt" kännetecken.
- Genetisk- beständigheten hos egenskaperna hos kromosomuppsättningen - karyotypen - och nukleotidsammansättningen av DNA hos individer av samma art. På grund av det faktum att icke-homologa kromosomer inte kan konjugera under meios, uppträder avkomman från korsningen av individer av olika arter med en ojämlik uppsättning kromosomer antingen inte alls eller är inte fertil. Detta skapar reproduktiv isolering av arten, upprätthåller dess integritet och säkerställer verkligheten av existens i naturen. Denna regel kan överträdas vid korsning av närbesläktade arter med samma karyotyp eller förekomst av olika mutationer, men undantaget bekräftar endast den allmänna regeln, och arten bör betraktas som stabila genetiska system. Det genetiska kriteriet är det huvudsakliga i systemet med artkriterier, men det är inte heller uttömmande.
Med all komplexitet i kriteriesystemet kan en art inte representeras som en grupp organismer som är absolut identiska i alla parametrar, det vill säga kloner. Tvärtom kännetecknas många arter av en betydande mängd jämna yttre tecken, som till exempel för vissa populationer av nyckelpigor är en dominans av röd färg karakteristisk, och för andra - svart.
Population är en strukturell enhet av en art och en elementär enhet av evolution
Det är svårt att föreställa sig att individer av samma art i verkligheten var jämnt fördelade över jordytan inom området, eftersom till exempel sjögrodan huvudsakligen lever i ganska sällsynta stillastående sötvattenförekomster, och det är osannolikt att det kan vara finns i åkrar och skogar. Arter i naturen faller oftast i separata grupper, beroende på de livsmiljöer som är lämpliga för kombinationen av förhållanden - befolkningen.
Befolkning- en grupp individer av samma art, som upptar en del av dess utbredningsområde, fritt förökar sig med varandra och relativt isolerade från andra populationer av individer av samma art under en mer eller mindre lång tid.
Populationer kan delas inte bara rumsligt, de kan till och med leva i samma territorium, men har skillnader i matpreferenser, häckningsdatum etc.
Således är en art en uppsättning populationer av individer som besitter ett antal gemensamma morfologiska, fysiologiska, biokemiska egenskaper och typer av relationer med miljön, som bor i ett visst område och även kan korsa sig med varandra för att bilda fertil avkomma, men nästan eller inte alls korsning med andra grupper, individer av samma art.
Inom arter med stora utbredningsområden, som täcker territorier med olika levnadsförhållanden, skiljer de sig ibland också åt underarter- stora populationer eller grupper av angränsande populationer med bestående morfologiska skillnader från andra populationer.
Populationer är inte utspridda över jordens yta slumpmässigt, de är bundna till specifika områden. Kombinationen av alla faktorer av livlös natur som är nödvändiga för bosättningen av individer av denna art kallas livsmiljö... Men bara dessa faktorer kanske inte räcker för att ockupera detta område av en population, eftersom den fortfarande måste vara involverad i nära interaktion med populationer av andra arter, det vill säga ta en viss plats i samhället av levande organismer - ekologisk nisch... Så den australiska pungdjurskoalabjörnen kan, allt annat lika, inte existera utan sin huvudsakliga födokälla - eukalyptus.
Populationer av olika arter som bildar en oskiljaktig enhet i samma livsmiljöer ger vanligtvis en mer eller mindre sluten cirkulation av ämnen och är elementära. ekologiska system(ekosystem) - biogeocenoser.
Trots all sin noggrannhet till miljöförhållanden är populationer av en art heterogena vad gäller yta, antal, täthet och rumslig fördelning av individer, och bildar ofta mindre grupper (familjer, flockar, besättningar, etc.), kön, ålder, genpool, etc. skiljer därför mellan deras storlek, ålder, kön, rumsliga, genetiska, etologiska och andra strukturer, såväl som dynamik.
Viktiga egenskaper hos befolkningen är genpool- en uppsättning gener som är karakteristiska för individer av en given population eller art, samt frekvensen av vissa alleler och genotyper. Olika populationer av samma art har till en början olika genpool, eftersom nya territorier utvecklas av individer med slumpmässiga och inte speciellt utvalda gener. Under påverkan av interna och externa faktorer genomgår genpoolen ännu mer betydande förändringar: den berikas på grund av uppkomsten av mutationer och en ny kombination av egenskaper och utarmas som ett resultat av förlusten av individuella alleler under döden eller migrationen av ett visst antal individer.
Nya egenskaper och deras kombinationer kan vara användbara, neutrala eller skadliga, därför är det bara individer som är anpassade till de givna miljöförhållandena som överlever och reproducerar sig framgångsrikt i befolkningen. Men på två olika punkter på jordens yta är miljöförhållandena aldrig helt identiska, därför kan förändringarnas riktning även i två närliggande populationer vara helt motsatta, eller så fortsätter de med olika hastighet... Resultatet av förändringar i genpoolen är divergensen av populationer i termer av morfologiska, fysiologiska, biokemiska och andra egenskaper. Om populationerna också är isolerade från varandra kan de ge upphov till nya arter.
Så uppkomsten av eventuella hinder vid korsningen av individer av olika populationer av samma art, till exempel på grund av bildandet av bergskedjor, förändringar i flodkanaler, skillnader i tidpunkten för reproduktion, etc., leder till det faktum att att populationer gradvis får fler och fler skillnader och så småningom blir olika arter. Under en tid, vid gränserna för dessa populationer, korsas individer och hybrider dyker upp, men med tiden försvinner dessa kontakter, det vill säga populationer från öppna genetiska system blir stängda.
Trots det faktum att miljöfaktorers verkan i första hand påverkar enskilda individer, är förändringen i den genetiska sammansättningen av en enskild organism obetydlig och kommer i bästa fall att manifestera sig endast i dess ättlingar. Underarter, arter och större taxa är inte heller lämpliga för rollen som elementära evolutionsenheter, eftersom de inte skiljer sig åt i morfologisk, fysiologisk, biokemisk, ekologisk, geografisk och genetisk enhet, medan populationer, som de minsta strukturella enheterna av en art, ackumulerar en mängd slumpmässiga förändringar, varav de värsta kommer att elimineras, motsvarar detta tillstånd och är evolutionens elementära enheter.
Mikroevolution
En förändring av populationernas genetiska struktur leder inte alltid till bildandet av en ny art, utan kan bara förbättra populationens anpassning till specifika miljöförhållanden, dock är arterna inte eviga och oförändrade - de kan utvecklas. Denna process av oåterkallelig historisk förändring av levande saker kallas evolution. Primära evolutionära transformationer sker inom arten på populationsnivå. De är först och främst baserade på mutationsprocessen och naturligt urval, vilket leder till en förändring i genpoolen av populationer och arten som helhet, eller till och med till bildandet av nya arter. Kombinationen av dessa elementära evolutionära händelser kallas mikroevolution.
Populationer kännetecknas av en enorm genetisk mångfald, som ofta inte manifesterar sig fenotypiskt. Genetisk mångfald uppstår från spontan mutagenes som sker kontinuerligt. De flesta mutationer är ogynnsamma för organismen och minskar livskraften för populationen som helhet, men om de är recessiva kan de finnas kvar i en heterozygot under lång tid. Vissa mutationer som inte har ett adaptivt värde under de givna existensförhållandena kan förvärva ett sådant värde i framtiden eller under utvecklingen av nya ekologiska nischer, vilket skapar en reserv av ärftlig variation.
Fluktuationer i antalet individer i populationer, migration och katastrofer, såväl som isolering av populationer och arter, har en betydande inverkan på mikroevolutionära processer.
En ny art är ett mellanresultat av evolutionen, men inte på något sätt dess resultat, eftersom detta inte avbryter mikroevolutionen - det fortsätter vidare. De framväxande nya arterna, i fallet med en framgångsrik kombination av egenskaper, befolkar nya livsmiljöer och ger i sin tur upphov till nya arter. Sådana grupper av närbesläktade arter kombineras till släkten, familjer etc. Evolutionära processer som förekommer i supraspecifika grupper kallas redan makroevolution. Till skillnad från makroevolution, mikroevolution fortskrider i en mycket kortare tidsram, medan den första kräver tiotals och hundratusentals och miljoner år, som till exempel mänsklig evolution.
Som ett resultat av mikroevolutionen bildas alla de olika arter av levande organismer som någonsin har funnits och som för närvarande lever på jorden.
Samtidigt är evolutionen oåterkallelig, och arter som redan har försvunnit dyker aldrig upp igen. De framväxande arterna konsoliderar allt som uppnåtts i evolutionsprocessen, men detta garanterar inte att det i framtiden inte kommer att dyka upp nya arter som kommer att ha bättre anpassningar till miljöförhållandena.
Bildande av nya arter
I vid bemärkelse innebär bildandet av nya arter inte bara avspaltning av en ny art från huvudstammen eller sönderfall av moderarten till flera dotter, utan också den allmänna utvecklingen av arten som ett integrerat system, vilket leder till till betydande förändringar i dess morfostrukturella organisation. Men oftare likadant artbildning betraktas som en process för att bilda nya arter genom förgrening av artens "släktträd".
En grundläggande lösning på problemet med artbildning föreslogs av Charles Darwin. Enligt hans teori leder spridningen av individer av en art till bildandet av populationer, som på grund av skillnader i miljöförhållanden tvingas anpassa sig till dem. Detta innebär i sin tur en förvärring av den intraspecifika kampen för tillvaron, styrd av naturligt urval. I dagsläget menar man att kampen för tillvaron inte alls är en obligatorisk faktor vid artbildning, tvärtom kan urvalstrycket i ett antal populationer minska. Skillnaden i existensvillkoren bidrar till uppkomsten av ojämlika adaptiva förändringar i artens populationer, vars konsekvens är divergensen mellan populationernas egenskaper och egenskaper - divergens.
Men ackumuleringen av skillnader, även på genetisk nivå, är inte på något sätt tillräcklig för att en ny art ska kunna uppstå. Så länge populationer som skiljer sig åt på något sätt inte bara är i kontakt, utan också kan föröka sig med bildandet av fertila avkommor, tillhör de samma art. Endast omöjligheten av flödet av gener från en grupp individer till en annan, även i händelse av förstörelse av de barriärer som skiljer dem åt, det vill säga korsning, innebär fullbordandet av den mest komplexa evolutionära processen för bildandet av en ny art .
Speciation är en fortsättning på mikroevolutionära processer. Det finns en synpunkt att artbildning inte kan reduceras till mikroevolution, den representerar ett kvalitativt evolutionsstadium och utförs genom andra mekanismer.
Specifikationsmetoder
Det finns två huvudsätt för artbildning: allopatrisk och sympatrisk.
Allopatrisk, eller geografisk artär en följd av den rumsliga separeringen av populationer genom fysiska barriärer (bergskedjor, hav och floder) på grund av deras uppkomst eller återbosättning i nya livsmiljöer (geografisk isolering). Eftersom genpoolen i den separerade populationen i det här fallet skiljer sig avsevärt från moderns, och förhållandena i dess livsmiljö inte kommer att sammanfalla med de ursprungliga, kommer detta med tiden att leda till divergens och bildandet av en ny art. Ett slående exempel på geografisk artbildning är den mångfald av finkarter som upptäcktes av Charles Darwin under sin resa på Beagleskeppet. Galapagosöarna nära Ecuadors kust. Tydligen hamnade enskilda individer av den enda fink som bor på den sydamerikanska kontinenten på något sätt på öarna, och på grund av skillnaderna i förhållanden (främst mattillgång) och geografisk isolering utvecklades de gradvis och bildade en grupp besläktade arter.
Vid hjärtat av sympatrisk, eller biologisk artbildning någon form av reproduktiv isolering ligger, med nya arter som dyker upp inom den ursprungliga artens räckvidd. En förutsättning för sympatrisk artbildning är den snabba isoleringen av de bildade formerna. Detta är en snabbare process än allopatrisk artbildning, och de nya formerna liknar sina ursprungliga förfäder.
Sympatisk artbildning kan orsakas av snabba förändringar i kromosomuppsättningen (polyploidisering) eller kromosomförändringar. Ibland uppstår nya arter genom hybridiseringen av två ursprungliga arter, till exempel tamplommon, som är en hybrid av svarttorn och körsbärsplommon. I vissa fall är sympatrisk artbildning associerad med separationen av ekologiska nischer i populationer av samma art inom ett enda område eller säsongsmässig isolering - skillnaden mellan tidpunkten för reproduktion i växter (olika tallarter i Kalifornien är dammiga i februari och april ) och tidpunkten för reproduktion hos djur.
Av all mångfald av nyuppkommande arter är det bara ett fåtal, de mest anpassade, som kan existera under lång tid och ge upphov till nya arter. Orsakerna till att de flesta arter dör är fortfarande okända, troligen beror detta på plötsliga klimatförändringar, geologiska processer och deras förskjutning av mer anpassade organismer. För närvarande är en av orsakerna till döden av ett betydande antal arter människan, som utrotar de största djuren och de vackraste växterna, och om på 1600-talet denna process precis hade börjat med utrotningen av den sista omgången, då under 2000-talet försvinner mer än 10 arter varje timme.
Bevarande av arternas mångfald som grund för biosfärens hållbarhet
Trots det faktum att det enligt olika uppskattningar finns 5-10 miljoner arter av organismer som ännu inte har beskrivits, kommer vi aldrig att veta om existensen av de flesta av dem, eftersom cirka 50 arter försvinner från jordens yta varje gång. timme. Försvinnandet av levande organismer för närvarande är inte nödvändigtvis förknippat med deras fysiska utrotning, oftare beror detta på förstörelse som ett resultat mänsklig aktivitet deras naturliga livsmiljöer. Det är osannolikt att en enskild art dör leder till ödesdigra konsekvenser för biosfären, men det är sedan länge fastställt att utrotningen av en växtart innebär att 10–12 djurarter dör, och detta utgör redan ett hot både för existensen. av enskilda biogeocenoser och för det globala ekosystemet som helhet.
De sorgliga fakta som samlats under de föregående decennierna tvingade Internationella unionen för bevarande av natur och naturresurser (IUCN) att börja samla in information om sällsynta och hotade växt- och djurarter 1949. 1966 publicerade IUCN den första Red Book of Facts.
röd bok- detta är officiellt dokument innehåller regelbundet uppdaterade uppgifter om status och utbredning av sällsynta och hotade arter av växter, djur och svampar.
Detta dokument antog en femstegsskala för statusen för en skyddad art, och det första skyddsskedet inkluderar arter vars räddning är omöjlig utan genomförande av särskilda åtgärder, och den femte - återställda arter, vars tillstånd beror på vidtagna åtgärder inte orsakar oro, men de är ännu inte föremål för industriell användning. Utvecklingen av en sådan skala gör det möjligt att rikta prioriterade insatser på skyddsområdet just till de mest sällsynta arterna, såsom Amurtigrarna.
Utöver den internationella versionen av Röda boken finns det även nationella och regionala versioner av den. I Sovjetunionen upprättades den röda boken 1974, och i Ryska federationen regleras förfarandet för att upprätthålla det av de federala lagarna "om miljöskydd", "om djurriket" och dekretet från Rysslands regering Federation "Om Ryska federationens röda bok". Idag finns 610 växtarter, 247 djurarter, 42 lavarter och 24 svamparter listade i Ryska federationens röda bok. Bestånden av några av dem, som en gång var utrotningshotade (europeisk bäver, europeisk bison), har redan framgångsrikt återställts.
Följande djurarter tas under skydd i Ryssland: rysk desman, tarbagan (mongoliskt murmeldjur), isbjörn, kaukasisk europeisk mink, havsutter, manul, Amur tiger, leopard, Snöleopard, sjölejon, valross, sälar, delfiner, valar, Przewalskis häst, kulan, rosa pelikan, vanlig flamingo, svart stork, liten svan, stäppörn, kungsörn, svart trana, sibirisk trana, bustard, örnuggla, vitmås, Medelhavet sköldpadda, japansk orm, gurza, vasspadda, kaspisk lampöga, alla typer av störfisk, sjölax, hjortbagge, ovanlig humla, vanlig apollo, mantisräka, pärlmussla, etc.
Växterna i den ryska federationens röda databok inkluderar 7 typer av snödroppar, vissa typer av malört, äkta ginseng, 7 typer av klockor, tandad ek, blånäbb, 11 arter av späckhuggare, rysk hasselripa, Schrencks tulpan, nöt- bärande lotus, Venus äkta toffel, finbladig pion, fjädergräs, Julias primula, lumbago (drömgräs) äng, belladonna belladonna, pitsunda tall, idegran, kinesisk vildblomma, lakustrin svamp, mjuk sphagnum, filiformophora, lockig phyllophora etc.
Sällsynta svampar representeras av sommartryffel, eller rysk svart tryffel, lackad tindersvamp, etc.
Skyddet av sällsynta arter är i de flesta fall förknippat med förbudet mot deras förstörelse, deras bevarande i en artificiellt skapad livsmiljö (zoo), skyddet av deras livsmiljöer och skapandet av genetiska banker med låg temperatur.
Mest effektiv åtgärd skydd av sällsynta arter är bevarandet av deras livsmiljöer, vilket uppnås genom att organisera ett nätverk av särskilt skyddade skyddade områden, som, i enlighet med den federala lagen "On Specially Protected Natural Areas" (1995), har internationella, federala, regionala eller lokal betydelse. Dessa inkluderar statliga naturreservat, nationalparker, naturparker, statliga naturreservat, naturminnen, dendrologiska parker, botaniska trädgårdar m.m.
Statens naturreservat- detta är ett särskilt skyddat naturkomplex (mark, vattendrag, mineraltillgångar, flora och fauna) helt tillbakadraget från ekonomisk användning, som har miljömässig, vetenskaplig, ekologisk och pedagogisk betydelse som ett exempel på den naturliga miljön, typiska eller sällsynta landskap, platser för bevarande av den genetiska fonden för växt- och djurvärlden.
Reserver som är en del av det internationella systemet av biosfärreservat, som genomför en global miljöövervakning, har status statliga naturliga biosfärområden... Reservatet är en naturvårds-, forsknings- och miljöutbildningsinstitution som syftar till att bevara och studera det naturliga förloppet av naturliga processer och fenomen, den genetiska fonden för flora och fauna, enskilda arter och samhällen av växter och djur, typiska och unika ekologiska system.
För närvarande finns det cirka 100 statliga naturreservat i Ryssland, varav 19 har status som biosfärreservat, inklusive Baikalsky, Barguzinsky, Caucasian, Kedrovaya Pad, Kronotsky, Prioksko-Terrasny, etc.
Till skillnad från reservat, territorier (vattenområden) National Parker omfattar naturkomplex och föremål av särskilt ekologiskt, historiskt och estetiskt värde, och är avsedda att användas i naturvårds-, utbildnings-, vetenskapliga och kulturella ändamål och för reglerad turism. Denna status har 39 speciellt skyddade naturområden, inklusive nationalparkerna Zabaikalsky och Sochi, såväl som nationalparkerna "Curonian Spit", "Russian North", "Shushensky Bor", etc.
Naturparkerär natunder jurisdiktionen för de ingående enheterna i Ryska federationen, vars territorier (vatten) inkluderar naturliga komplex och föremål av betydande ekologiskt och estetiskt värde och är avsedda för användning i naturskydd, utbildnings- och rekreationsändamål.
Statens naturreservatär territorier (vattenområden) som är av särskild betydelse för bevarandet eller återställandet av naturliga komplex eller deras komponenter och upprätthållandet av den ekologiska balansen.
Utveckling av evolutionära idéer. Betydelsen av evolutionsteorin om Charles Darwin. Sammankopplingen av evolutionens drivkrafter. Former av naturligt urval, typer av kamp för tillvaron. Syntetisk evolutionsteori. Elementära evolutionsfaktorer. Forskning av S. S. Chetverikov. Evolutionsteorins roll i bildandet av den moderna naturvetenskapliga bilden av världen
Utveckling av evolutionära idéer
Alla teorier om ursprunget och utvecklingen av den organiska världen kan reduceras till tre huvudriktningar: kreationism, transformationism och evolutionism. Kreationismär begreppet artbeständighet, med tanke på mångfalden av den organiska världen som ett resultat av dess skapelse av Gud. Denna trend bildades som ett resultat av etableringen av den kristna kyrkans dominans i Europa, baserad på bibliska texter. Framstående företrädare för kreationismen var C. Linnaeus och J. Cuvier.
"Botanikernas prins" K. Linnaeus, som upptäckte och beskrev hundratals nya växtarter, och skapade deras första harmoniska system, bevisade ändå att det totala antalet arter av organismer har förblivit oförändrat sedan jordens skapelse, dvs. de inte bara dyker inte upp igen, utan försvinner inte. Först mot slutet av sitt liv kom han till slutsatsen att Guds händers verk är förlossning, medan arter kan utvecklas som ett resultat av anpassning till lokala förhållanden.
Den framstående franske zoologen J. Cuviers (1769-1832) bidrag till biologin baserades på många data från paleontologi, jämförande anatomi och fysiologi teori om korrelationer- förhållandet mellan delar av kroppen. Tack vare detta blev det möjligt att rekonstruera djurets yttre utseende i separata delar. Men i processen av paleontologisk forskning kunde J. Cuvier inte låta bli att uppmärksamma både det skenbara överflöd av fossila former och de skarpa förändringarna i djurgrupper under geologisk historia. Dessa data tjänade som utgångspunkt för formuleringen katastrofteori, enligt vilken alla eller nästan alla organismer på jorden upprepade gånger har dött till följd av periodiska naturkatastrofer, och sedan återkoloniserades planeten av arter som överlevde katastrofen. Anhängarna av J. Cuvier räknade upp till 27 sådana katastrofer i jordens historia. Men överväganden om evolution tycktes J. Cuvier vara skild från verkligheten.
Motsättningarna i kreationismens initiala positioner, som blev mer och mer uppenbara i takt med ackumuleringen av vetenskapliga fakta, fungerade som utgångspunkt för bildandet av ett annat synssystem - transformism, som erkänner arternas verkliga existens och deras historiska utveckling. Representanter för denna trend - J. Buffon, I. Goethe, E. Darwin och E. Geoffroy Saint-Hilaire, som inte kunde avslöja de verkliga orsakerna till evolutionen, reducerade dem till anpassning till miljöförhållanden och arvet av förvärvade karaktärer. Transformismens rötter kan hittas i verk av antika grekiska och medeltida filosofer som erkände de historiska förändringarna i den organiska världen. Således uttryckte Aristoteles idén om naturens enhet och den gradvisa övergången från kroppar av livlös natur till växter och från dem till djur - "naturens stege". Han ansåg att huvudorsaken till förändringarna i levande organismer var deras inre strävan efter perfektion.
Den franske naturforskaren J. Buffon (1707-1788), vars huvudsakliga livsverk var 36-volymen "Naturhistoria", i motsats till kreationisternas idéer, utökade ramen för jordens historia till 80-90 tusen år. Samtidigt noterade han enheten i floran och faunan, såväl som möjligheten att förändra relaterade organismer under påverkan av miljöfaktorer som ett resultat av domesticering och hybridisering.
Den engelske läkaren, filosofen och poeten E. Darwin (1731-1802), Charles Darwins farfar, trodde att den organiska världens historia är miljontals år gammal, och mångfalden i djurvärlden är resultatet av att blanda flera "naturliga" grupper, inflytande yttre miljön, träning och icke-träning av organ och andra faktorer.
Enheten i planen för strukturen av grupper av djur ansågs vara ett av de viktigaste bevisen för utvecklingen av den levande världen av E. Geoffroy Saint-Hilaire (1772–1844). Men till skillnad från sina föregångare var han benägen att tro att förändringen i arter beror på påverkan av miljöfaktorer inte på vuxna, utan på embryona.
Trots att i diskussionen som 1831 blossade upp mellan J. Cuvier och E. Geoffroy Saint-Hilaire i form av en serie reportage vid Vetenskapsakademien, den tydliga övervikten kvarstod på den förres sida, var det transformism som blev evolutionismens föregångare. Evolutionism(evolutionsteori, evolutionär lära) är ett trossystem som erkänner naturens utveckling enligt vissa lagar. Det är biologins teoretiska höjdpunkt, som gör att vi kan förklara mångfalden och komplexiteten hos levande system vi observerar. Men på grund av det faktum att den evolutionära doktrinen beskriver fenomen som är svåra att observera, stöter den på betydande svårigheter. Ibland kallas evolutionsteorin "darwinism" och identifieras med Charles Darwins läror, vilket är fundamentalt felaktigt, för även om Charles Darwins teori gjorde ett ovärderligt bidrag till utvecklingen av inte bara evolutionär lära, utan biologi i allmänhet (liksom många andra vetenskaper), grunden för evolutionsteorin lades av andra vetenskapsmän, den fortsätter att utvecklas till denna dag, och "darwinism" har i många aspekter endast historisk betydelse.
Skaparen av den första evolutionsteorin - lamarckismen - var den franske naturforskaren JB Lamarck (1744–1829). Han ansåg att evolutionens drivkraft var organismernas inre strävan efter perfektion ( graderingslagen), men anpassning till miljöförhållanden tvingar dem att avvika från denna huvudlinje. Samtidigt utvecklas de organ som används intensivt av djuret i livets process och är onödiga för honom, tvärtom, försvagas och kan till och med försvinna ( lagen om motion och icke-träning av organ). De tecken som förvärvats i livets process fixeras och förs vidare till ättlingar. Så, närvaron av membran mellan tårna på vattenfåglar, förklarade han med försöken från deras förfäder att röra sig i vattenmiljön, och giraffernas långa hals, enligt Lamarck, är en konsekvens av det faktum att deras förfäder försökte få löv från trädtopparna.
Nackdelarna med lamarckismen var den teoretiska karaktären hos många konstruktioner, såväl som medgivandet av Skaparens ingripande i evolutionen. I processen för utveckling av biologi blev det klart att individuella förändringar som förvärvats av organismer i livets process, för det mesta, passar inom gränserna för fenotypisk variabilitet, och deras överföring är praktiskt taget omöjlig. Till exempel skar den tyske zoologen och evolutionsläran A. Weismann (1834–1914) svansen av möss i många generationer och fick alltid bara svansgnagare i avkomman. J. B. Lamarcks teori accepterades inte av hans samtid, men vid sekelskiftet låg den till grund för det s.k. nylamarckism.
Betydelsen av evolutionsteorin om Charles Darwin
Förutsättningarna för skapandet av Charles Darwins mest kända evolutionsteorin, eller darwinismen, var publiceringen 1778 av den engelska ekonomen T. Malthus verk "A Treatise on Population", geologen Charles Lyells verk, formuleringen. av cellteorin, framgången med urval i England och Ch. Darwin (1809-1882), som gjordes under studieåren vid Cambridge, på expeditionen som naturforskare på skeppet "Beagle" och efter dess färdigställande.
Så, T. Malthus hävdade att jordens befolkning ökar exponentiellt, vilket avsevärt överstiger planetens förmåga att förse den med mat och leder till att några av avkomman dör. De paralleller som drogs av Charles Darwin och hans medförfattare A. Wallace (1823–1913) visade att individer i naturen reproducerar sig i mycket hög hastighet, men populationsstorleken förblir relativt konstant. Den engelske geologen C. Lyells studier gjorde det möjligt att fastställa att jordens yta långt ifrån alltid var densamma som den är nu, och dess förändringar orsakades av effekterna av vatten, vind, vulkanutbrott och aktiviteten hos levande organismer. Charles Darwin själv, även under sina studentår, slogs av skalbaggarnas extrema variationsgrad och under resan - av likheten mellan floran och faunan på det kontinentala Sydamerika och de närliggande Galapagosöarna, och samtidigt en betydande mångfald av arter, till exempel finkar och sköldpaddor. Dessutom kunde han på expeditionen observera skelett av gigantiska utdöda däggdjur, liknande moderna bältdjur och sengångare, vilket avsevärt skakade hans tro på skapandet av arter.
De viktigaste bestämmelserna i evolutionsteorin uttrycktes av Charles Darwin 1859 vid ett möte i Royal Society of London och utvecklades senare i böckerna The Origin of Species by Natural Selection, or the Preservation of Favoured Breeds in the Struggle for Life (1859), Föränderliga husdjur och odlade växter"(1868)," Människans ursprung och sexuellt urval "(1871)," uttryck för känslor hos människor och djur "(1872), etc.
Essence utvecklad av Charles Darwin evolutionsbegrepp kan reduceras till ett antal följdbestämmelser som har lämpliga bevis:
- Individer som utgör vilken population som helst producerar många fler avkommor än vad som är nödvändigt för att behålla populationens storlek.
- På grund av det faktum att de vitala resurserna för alla slags levande organismer är begränsade, uppstår oundvikligen mellan dem kamp för tillvaron... C. Darwin skiljde mellan intraspecifik och interspecifik kamp, såväl som kamp med miljöfaktorer. Samtidigt påpekade han också att vi inte bara talar om en viss individs kamp för tillvaron, utan också för att överge avkomma.
- Konsekvensen av kampen för tillvaron är naturligt urval- rådande överlevnad och reproduktion av organismer som råkar vara mest anpassade till de givna existensvillkoren. Naturligt urval är på många sätt analogt med artificiellt urval, som människan har använt sedan urminnes tider för att utveckla nya varianter av växter och raser av husdjur. Genom att välja individer som har någon önskvärd egenskap, behåller en person dessa egenskaper genom artificiell avel genom selektiv avel eller pollinering. En speciell form av naturligt urval är sexuellt urval för egenskaper som vanligtvis inte har en direkt adaptiv betydelse (långa fjädrar, enorma horn, etc.), men bidrar till att uppnå reproduktiv framgång, eftersom de gör en individ mer attraktiv för motsatsen. kön eller mer formidabel för rivaler av samma kön.
- Materialet för evolutionen är de skillnader i organismer som uppstår till följd av deras variabilitet. C. Darwin skiljde mellan obestämd och bestämd variabilitet. Vissa(grupp)variabilitet visar sig hos alla individer av arten på samma sätt under påverkan av en viss faktor och försvinner hos avkomman när effekten av denna faktor upphör. Obestämd(individuell) variabilitet är de förändringar som sker hos varje enskild individ, oavsett fluktuationer i miljöfaktorernas värden, och som överförs till ättlingar. Sådan variation har inte en adaptiv (adaptiv) karaktär. Därefter visade det sig att en viss variation är icke-ärftlig och en obestämd är ärftlig.
- Naturligt urval leder i slutändan till en divergens i egenskaperna hos enskilda isolerade arter - divergens, och i slutändan till bildandet av nya arter.
Charles Darwins evolutionsteori postulerade inte bara processen för arters uppkomst och utveckling, utan avslöjade också själva mekanismen för evolution, som är baserad på principen om naturligt urval. Darwinismen förnekade också att evolutionen var programmerad och postulerade dess kontinuerliga natur.
Samtidigt kunde inte Charles Darwins evolutionsteori svara på ett antal frågor, till exempel om genetiskt materials natur och dess egenskaper, essensen av ärftlig och icke-ärftlig variation, deras evolutionära roll. Detta ledde till darwinismens kris och uppkomsten av nya teorier: neo-lamarckism, saltationism, begreppet nomogenes, etc. Nylamarckism baserad på J. B. Lamarcks teori om arvet av förvärvade karaktärer. Saltationismär ett system av åsikter om evolutionsprocessen som abrupta förändringar som leder till att nya arter, släkten och större systematiska grupper uppstår snabbt. Begrepp nomogenes postulerar den programmerade riktningen för evolutionen och utvecklingen av olika tecken på grundval av interna lagar. Endast syntesen av darwinism och genetik på 1920- och 1930-talen kunde övervinna de motsättningar som oundvikligen uppstod när man förklarade ett antal fakta.
Sammankopplingen av evolutionens drivkrafter
Evolution kan inte associeras med verkan av någon faktor, eftersom mutationer i sig är slumpmässiga och oriktade förändringar, och inte kan ge individers anpassning till miljöfaktorer, medan naturligt urval redan sorterar dessa förändringar. På samma sätt kan selektion i sig inte vara den enda faktorn i evolutionen, eftersom selektion kräver lämpligt material som tillhandahålls av mutationer.
Det kan dock noteras att mutation och genflöde skapar variabilitet, medan naturligt urval och gendrift sorterar denna variabilitet. Detta innebär att de faktorer som skapar variabilitet startar mikroevolutionsprocessen, medan de som sorterar variabilitet fortsätter den, vilket leder till etableringen av nya frekvenser av varianter. Således kan evolutionära förändringar inom en population ses som ett resultat av verkan av motsatta krafter som skapar och sorterar genotypisk variabilitet.
Ett exempel på interaktionen mellan mutation och selektion är hemofili hos människor. Blödarsjuka är ett tillstånd som orsakas av minskad blodpropp. Tidigare ledde det till döden under den pre-reproduktiva perioden, eftersom skador på mjukvävnader potentiellt kan leda till stor blodförlust. Denna sjukdom orsakas av en recessiv mutation i den könsbundna H (Xh)-genen. Kvinnor får sällan hemofili, oftare är de heterozygota bärare, men deras söner kan ärva denna sjukdom. Teoretiskt sett, under flera generationer, dör sådana män före puberteten och gradvis bör denna allel försvinna från befolkningen, dock minskar inte förekomsten av denna sjukdom på grund av upprepade mutationer i detta locus, som hände i drottning Victoria, som överförde sjukdomen till tre generationer av Europas kungahus. Den konstanta förekomsten av denna sjukdom indikerar en balans mellan mutationsprocessen och urvalstrycket.
Former av naturligt urval, typer av kamp för tillvaron
Naturligt urval de kallar selektiv erfarenhet och övergivande av avkomma av de mest vältränade individerna och döden av de minst vältränade.
Kärnan i naturligt urval i evolutionsteorin ligger i det differentierade (icke-slumpmässiga) bevarandet av vissa genotyper i befolkningen och deras selektiva deltagande i överföringen av gener till nästa generation. Dessutom påverkar det inte en enda egenskap (eller gen), utan hela fenotypen, som bildas som ett resultat av genotypens interaktion med miljöfaktorer. Naturligt urval kommer inte att vara detsamma under olika miljöförhållanden. För närvarande särskiljs flera former av naturligt urval: stabiliserande, drivande och störande.

Stabiliserande urval syftar till att konsolidera en snäv reaktionshastighet, som visade sig vara mest gynnsam under de givna existensförhållandena. Det är typiskt för de fall då fenotypiska egenskaper är optimala för oföränderliga miljöförhållanden. Ett slående exempel på effekten av stabiliserande urval är upprätthållandet av en relativt konstant kroppstemperatur hos varmblodiga djur. Denna form av urval studerades i detalj av den enastående ryske zoologen I.I.Shmalgauzen.
Val av körning uppstår som svar på förändringar i miljöförhållanden, som ett resultat av vilka mutationer som avviker från egenskapens genomsnittliga värde kvarstår, medan den tidigare dominerande formen är föremål för förstörelse, eftersom den inte tillräckligt uppfyller de nya existensvillkoren. Till exempel, i England, som ett resultat av luftföroreningar från industriella utsläpp, var björkfjärilar med mörkfärgade vingar, som var mindre märkbara för fåglar mot bakgrund av rökta björkstammar, utbredda hittills osynliga på många ställen. Val av körning bidrar inte till att fullständigt eliminera formen mot vilken den agerar, eftersom, på grund av åtgärder som vidtagits av regeringen och miljöorganisationer, situationen med luftföroreningar har förbättrats dramatiskt, och färgen på fjärilen återkommer. vingar till originalversionen.
Slitande, eller störande urval gynnar bevarandet av extrema varianter av egenskapen och tar bort intermediära sådana, som till exempel, som ett resultat av användningen av bekämpningsmedel, uppstår grupper av insekter som är resistenta mot det. Genom sin mekanism är disruptivt urval motsatsen till stabiliserande urval. På grund av denna form av selektion uppstår flera kraftigt differentierade fenotyper i befolkningen. Detta fenomen kallas polymorfism... Uppkomsten av reproduktiv isolering mellan skarpt uttryckta former kan leda till artbildning.
Ibland övervägs de också separat destabiliserande urval, som behåller mutationer som leder till en mängd olika egenskaper, till exempel färgen och strukturen på skalen hos vissa blötdjur som lever i de heterogena mikroförhållandena i havets steniga surfremsa. Denna form av urval upptäcktes av D.K.Belyaev i studien av domesticering av djur.
I naturen finns ingen av formerna av naturligt urval i sin rena form, utan tvärtom finns det olika kombinationer av dem, och när miljöförhållandena förändras kommer den ena eller den andra av dem i förgrunden. Så när förändringar i miljön har genomförts ersätts körvalet med stabiliserande urval, vilket optimerar gruppen av individer i de nya existensvillkoren.
Naturligt urval sker på olika nivåer, och skiljer därför också på individuellt, grupp- och sexuellt urval. Enskild selektion eliminerar mindre anpassade individer från deltagande i reproduktion, medan gruppselektion syftar till att bevara en egenskap som inte är användbar för en enskild individ, utan för gruppen som helhet. Under press grupp selektion kan helt dö ut hela populationer, arter och större grupper av organismer utan att lämna avkomma. Till skillnad från individuellt urval minskar gruppval variationen av former i naturen.
Sexuellt urval utförs inom en våning. Det främjar utvecklingen av egenskaper som säkerställer framgång med att lämna den största avkomman. Tack vare denna form av naturligt urval har sexuell dimorfism utvecklats, vilket uttrycks i storleken och färgen på påfågelns svans, hjorthorn m.m.
Naturligt urval är resultatet kamp för tillvaron baserat på ärftlig variation. Kampen för tillvaron förstås som hela uppsättningen av relationer mellan individer av deras egen och andra arter, såväl som med abiotiska miljöfaktorer. Detta förhållande avgör framgången eller misslyckandet för en viss individ i överlevnad och fortplantning. Anledningen till kampen för tillvaron är uppkomsten av ett överskott av individer i förhållande till tillgängliga resurser. Utöver konkurrens bör dessa relationer innefatta ömsesidig hjälp, vilket ökar individernas chanser att överleva.
Interaktion med miljöfaktorer kan också leda till att de allra flesta individer dör, till exempel hos insekter, av vilka endast en liten del överlever vintern.
Syntetisk evolutionsteori
Genetikens framgångar i början av 1900-talet, till exempel upptäckten av mutationer, antydde att ärftliga förändringar i organismers fenotyp inträffar plötsligt och inte bildas på länge, vilket postulerades av Charles evolutionsteorin Darwin. Men ytterligare forskning inom området för populationsgenetik ledde till formuleringen på 20-50-talet av 1900-talet, ett nytt system av evolutionära åsikter - syntetisk evolutionsteori... Ett betydande bidrag till dess skapelse gjordes av forskare från olika länder: sovjetiska forskare S.S.Chetverikov, I.I.Shmalgauzen och A.N.Severtsov, engelsk biokemist och genetiker D. Haldane, amerikanska genetiker S. Huxley, paleontolog D. Simpson och zoolog E. Mayr.
De viktigaste bestämmelserna i den syntetiska evolutionsteorin:
- Evolutionens elementära material är ärftlig variation (mutationell och kombinativ) hos individer i befolkningen.
- Evolutionens elementära enhet är en befolkning där alla evolutionära förändringar äger rum.
- Ett elementärt evolutionärt fenomen är en förändring i den genetiska strukturen hos en population.
- Elementära evolutionsfaktorer - gendrift, livsvågor, genflöde - är av oriktad, slumpmässig natur.
- Den enda riktningsfaktorn i evolutionen är naturligt urval, vilket är kreativt. Naturligt urval är stabiliserande, framdrivande och störande.
- Evolutionen är divergent, det vill säga ett taxon kan ge upphov till flera nya taxa, medan varje art bara har en förfader (art, population).
- Evolutionen är gradvis och kontinuerlig. Speciation som ett stadium i den evolutionära processen är en sekventiell ersättning av en population med en följd av andra tillfälliga populationer.
- Det finns två typer av evolutionära processer: mikroevolution och makroevolution. Makroevolution har inga egna speciella mekanismer och utförs endast tack vare mikroevolutionära mekanismer.
- Vilken systematisk grupp som helst kan antingen frodas (biologiska framsteg) eller dö ut (biologisk regression). Biologiska framsteg uppnås genom förändringar i organismers struktur: aromorfoser, idioadaptationer eller allmän degeneration.
- Evolutionens huvudsakliga lagar är dess oåterkalleliga natur, den progressiva komplikationen av livsformer och utvecklingen av arternas anpassning till miljön. Det har dock inte evolutionen ultimat mål, det vill säga processen är oriktad.
Trots att evolutionsteorin under de senaste decennierna har berikats med data från relaterade vetenskaper - genetik, avel, etc., tar den fortfarande inte hänsyn till ett antal aspekter, till exempel en riktad förändring av arvsmaterialet, därför är det i framtiden möjligt att skapa ett nytt evolutionsbegrepp som kommer att ersätta den syntetiska teorin. ...
Elementära evolutionsfaktorer
Enligt den syntetiska evolutionsteorin består ett elementärt evolutionärt fenomen i en förändring av den genetiska sammansättningen av en population, och händelser och processer som leder till en förändring i genpooler kallas elementära evolutionsfaktorer... Dessa inkluderar mutation, befolkningsvågor, gendrift, isolering och naturligt urval. Med tanke på den exceptionella betydelsen av naturligt urval i evolutionen kommer det att övervägas separat.
Mutationsprocess som är lika kontinuerlig som själva evolutionen, upprätthåller den genetiska heterogeniteten i befolkningen på grund av uppkomsten av fler och fler nya varianter av gener. Mutationer som uppstår under påverkan av yttre och inre faktorer klassificeras som genetiska, kromosomala och genomiska.
Genmutationer förekommer med en frekvens av 10 –4 –10 –7 per gamet, men eftersom det totala antalet gener hos människor och de flesta högre organismer kan nå flera tiotusentals är det omöjligt att föreställa sig att två organismer skulle vara helt identiska. De flesta av de mutationer som uppstår är recessiva, speciellt eftersom de dominerande mutationerna omedelbart genomgår naturligt urval. Recessiva mutationer skapar själva reserven av ärftlig variabilitet, men innan de uppträder i fenotypen måste de fixeras hos många individer i ett heterozygott tillstånd på grund av fri korsning i populationen.
Kromosomala mutationer associerade med förlust eller överföring av en del av en kromosom (en hel kromosom) till en annan, är också ganska vanliga i olika organismer, till exempel ligger skillnaden mellan vissa råttarter i ett enda kromosompar, vilket gör det svårt att korsa dem.
Genomiska mutationer associerade med polyploidisering, leder också till reproduktiv isolering av den nyuppkomna populationen på grund av störningar i mitos av den första divisionen av zygoten. I växter är de dock ganska utbredda och sådana växter kan växa i Arktis och på alpina ängar på grund av deras större motståndskraft mot miljöfaktorer.
Kombinativ variabilitet, som säkerställer uppkomsten av nya varianter av att kombinera gener i genotypen, och följaktligen ökar sannolikheten för uppkomsten av nya fenotyper, bidrar också till evolutionära processer, eftersom endast hos människor är antalet varianter av kromosomkombinationer 2 23, det vill säga utseendet på en organism som liknar befintliga, är det nästan omöjligt.
Befolkningsvågor. Det motsatta resultatet (utarmning av den genetiska sammansättningen) orsakas ofta av fluktuationer i antalet organismer i naturliga populationer, som hos vissa arter (insekter, fiskar etc.) kan förändras tiotals och hundratals gånger - befolkningsvågor, eller "Livets vågor"... En ökning eller minskning av antalet individer i populationer kan vara som följer periodisk och icke periodisk... De förstnämnda är säsongsbetonade eller fleråriga, såsom flyttfåglar eller häckning i Daphnia, där det på våren och sommaren bara finns honor, och på hösten verkar hanarna vara nödvändiga för sexuell reproduktion. Icke-periodiska fluktuationer i antal orsakas ofta av en kraftig ökning av mängden mat under ett gynnsamt år, brott mot livsmiljöförhållanden, reproduktion av skadedjur eller rovdjur.
Eftersom återställandet av befolkningens storlek sker på bekostnad av inte ett stort antal individer som inte har hela uppsättningen av alleler, kommer de nya och ursprungliga populationerna att ha en annan genetisk struktur. Förändringen i frekvensen av gener i en population under påverkan av slumpmässiga faktorer kallas gendrift, eller genetiska automatiska processer... Det sker också under utvecklingen av nya territorier, eftersom ett extremt begränsat antal individer av en given art faller på dem, vilket kan ge upphov till en ny population. Därför kan dessa individers genotyper ( grundareffekt). Som ett resultat av gendrift klyvs ofta nya homozygota former (för mutanta alleler) ut, vilket kan vara adaptivt värdefullt och sedan kommer att plockas upp av naturligt urval.
Så bland den indiska befolkningen på den amerikanska kontinenten och lappländarna finns det en mycket hög andel personer med I (0) blodgrupp, medan III och IV grupper är extremt sällsynta. Förmodligen, i det första fallet, var befolkningens grundare personer som inte hade I B-allelen, eller så gick den förlorad i urvalsprocessen.
Fram till en viss punkt sker ett utbyte av alleler mellan närliggande populationer som ett resultat av korsning mellan individer av olika populationer - genflöde, vilket minskar diskrepansen mellan individuella populationer, men det slutar med början av isolering. I huvudsak är genflödet en fördröjd mutationsprocess.
Isolering. Eventuella förändringar i populationens genetiska struktur måste fixas, vilket beror på isolering- uppkomsten av eventuella hinder (geografiska, ekologiska, beteendemässiga, reproduktiva, etc.) som komplicerar och gör det omöjligt för individer av olika populationer att passera. Även om isolering i sig inte skapar nya former, bevarar den inte desto mindre genetiska skillnader mellan populationer som utsätts för naturligt urval. Det finns två former av isolering: geografisk och biologisk.
Geografisk isolering uppstår som ett resultat av uppdelningen av området med fysiska barriärer (vattenhinder för landlevande organismer, landområden för vattenlevande arter, alternerande upphöjda områden och slätter); detta underlättas av en stillasittande eller fast (i växter) livsstil. Ibland kan geografisk isolering orsakas av utvidgningen av en arts utbredningsområde, följt av utrotningen av dess populationer i de mellanliggande territorierna.
Biologisk isoleringär en konsekvens av vissa skillnader mellan organismer inom samma art, som på något sätt förhindrar fri korsning. Det finns flera typer av biologisk isolering: ekologisk, säsongsbetonad, etologisk, morfologisk och genetisk. Miljöisolering uppnås genom separation av ekologiska nischer (till exempel preferensen för vissa livsmiljöer eller matens natur, som i grankorsnäbb och tallkorsnäbb). Säsong(tillfällig) isolering observeras vid reproduktion av individer av samma art vid olika tidpunkter (olika sillflockar). Etologisk isolering beror på beteendets egenskaper (drag av uppvaktningsritualen, färgning, "sång" av kvinnor och män från olika populationer). På morfologisk isolering ett hinder för korsning är skillnaden i strukturen hos reproduktionsorganen eller till och med storleken på kroppen (Pekingese och Grand Danois). Genetisk isolering har störst inverkan och visar sig i inkompatibiliteten hos könsceller (zygotens död efter befruktning), sterilitet eller minskad livsduglighet hos hybrider. Skälen till detta är särdragen hos kromosomernas antal och form, som ett resultat av vilket det blir omöjligt att slutföra celldelning (mitos och meios).
Genom att störa fri korsning mellan populationer förstärker isoleringen därigenom i dem de skillnader som har uppstått på genotypnivå på grund av mutationer och fluktuationer i antal. Dessutom utsätts var och en av populationerna för naturligt urval separat från den andra, och detta leder i slutändan till divergens.
Det naturliga urvalets kreativa roll i evolutionen
Naturligt urval utför funktionen av ett slags "sil", sorterar genotyper efter graden av kondition. Charles Darwin betonade dock att urval inte bara och inte så mycket syftar till att bevara det exceptionellt bästa, utan till att ta bort det sämsta, det vill säga det låter dig bevara multivarians. Funktionen av naturligt urval är inte begränsad till detta, eftersom det säkerställer reproduktionen av anpassade genotyper, och därmed bestämmer riktningen för utvecklingen, successivt summerar slumpmässiga och många avvikelser. Naturligt urval har inget bestämt mål: på basis av samma material (ärftlig variation) under olika förhållanden kan olika resultat erhållas.
I detta avseende kan den aktuella evolutionsfaktorn inte jämföras med en skulptörs arbete som skär ett marmorblock, snarare fungerar han som en avlägsen förfader till människan och gör ett arbetsredskap från ett stenfragment, utan att föreställa sig det slutliga resultatet, vilket beror inte bara på stenens beskaffenhet och dess form, utan och på styrkan, slagets riktning etc. Men i händelse av misslyckande avvisar urval, som en humanoid varelse, den "oregelbundna" formen.
Urvalsavgift är förekomsten genetisk belastning, det vill säga ackumuleringen av mutationer i befolkningen, som med tiden kan bli dominerande på grund av att de flesta individer plötsligt dör eller migrationen av ett litet antal av dem.
Under trycket av naturligt urval bildas inte bara mångfalden av arter, utan deras organisationsnivå ökar också, inklusive deras komplikation eller specialisering. Men till skillnad från artificiellt urval som utförs av en person endast för ekonomiskt värdefulla egenskaper, ofta till nackdel för adaptiva egenskaper, kan det naturliga urvalet inte bidra till detta, eftersom ingen anpassning i naturen kan kompensera för skadan från en minskning av livskraften hos en befolkning .
Forskning S. S. Chetverikov
Ett av de viktiga stegen mot försoning av darwinism och genetik togs av Moskva-zoologen S. S. Chetverikov (1880-1959). Baserat på resultaten av att studera den genetiska sammansättningen av naturliga populationer av fruktflugan Drosophila, bevisade han att de bär på många recessiva mutationer i en heterozygot form som inte bryter mot den fenotypiska enhetligheten. De flesta av dessa mutationer är ogynnsamma för kroppen och skapar den sk genetisk belastning, vilket minskar befolkningens lämplighet för miljön. Vissa mutationer som inte har en adaptiv betydelse vid ett givet tillfälle i utvecklingen av en art kan få ett visst värde senare, och är därför reserv av ärftlig variation. Spridningen av sådana mutationer bland individer av befolkningen på grund av successiva fria korsningar kan i slutändan leda till deras övergång till ett homozygott tillstånd och manifestation i en fenotyp. Om det givna tillståndet för funktionen är hårtork- är adaptiv, så kommer den efter några generationer att helt förskjuta den dominerande fenen, som är mindre lämplig för dessa förhållanden, från befolkningen, tillsammans med dess bärare. På grund av sådana evolutionära förändringar bibehålls således endast den recessiva mutanta allelen, och dess dominanta allel försvinner.
Låt oss försöka bevisa detta med ett specifikt exempel. När man studerar en viss population kan det konstateras att inte bara dess fenotypiska, utan också genotypiska strukturer kan förbli oförändrade under lång tid, vilket beror på fri korsning, eller panmixia diploida organismer.
Detta fenomen beskrivs i lag Hardy - Weinberg, enligt vilken i en ideal population av oändligt stora storlekar i frånvaro av mutationer, migrationer, befolkningsvågor, gendrift, naturligt urval och under förutsättning av fri korsning, kommer frekvenserna av alleler och genotyper av diploida organismer inte att förändras under en antal generationer.
Till exempel, i en population kodas en viss egenskap av två alleler av samma gen - den dominanta ( A) och recessiv ( a). Frekvensen av den dominanta allelen betecknas som R och recessiv - q... Summan av frekvenserna för dessa alleler är 1: sid + q= 1. Därför, om vi vet frekvensen för den dominanta allelen, så kan vi bestämma frekvensen för den recessiva allelen: q = 1 – sid... Faktum är att allelfrekvenserna är lika med sannolikheterna för motsvarande könsceller. Sedan, efter bildandet av zygoter, kommer frekvenserna av genotyper redan i den första generationen att vara:
(pA + qa) 2 = sid 2 AA + 2pqAa + q 2 aa = 1,
var sid 2 AA- frekvens av dominanta homozygoter;
2pqAa- frekvens av heterozygoter;
q 2 aa- frekvensen av recessiva homozygoter.
Det är lätt att beräkna att i efterföljande generationer kommer frekvenserna av genotyper att förbli desamma, vilket upprätthåller den genetiska mångfalden i populationen. Men i naturen saknas idealpopulationer, och därför kan muterade alleler i dem inte bara bestå, utan också spridas och till och med ersätta tidigare vanligare alleler.
S.S.Chetverikov insåg tydligt att naturligt urval inte bara eliminerar individuella mindre framgångsrika egenskaper, och följaktligen de alleler som kodar för dem, utan också verkar på hela komplexet av gener som påverkar manifestationen av en viss gen i fenotypen, eller genotypisk miljö... Som en genotypisk miljö betraktas hela genotypen för närvarande som en uppsättning gener som kan förstärka eller försvaga manifestationen av specifika alleler.
Inte mindre viktigt i utvecklingen av evolutionär doktrin och forskning av S. S. Chetverikov inom området för befolkningsdynamik, i synnerhet "livsvågor" eller befolkningsvågor. Medan han fortfarande var student publicerade han 1905 en artikel om möjligheten av utbrott av massreproduktion av insekter och en lika snabb minskning av deras antal.
Evolutionsteorins roll i bildandet av den moderna naturvetenskapliga bilden av världen
Evolutionsteorins betydelse för utvecklingen av biologi och andra naturvetenskaper kan knappast överskattas, eftersom den var den första som förklarade villkoren, orsakerna, mekanismerna och resultaten av den historiska utvecklingen av livet på vår planet, det vill säga den gav en materialistisk förklaring av utvecklingen av den organiska världen. Dessutom var teorin om naturligt urval den första verkligt vetenskapliga teorin om biologisk evolution, eftersom Charles Darwin, när han skapade den, inte förlitade sig på spekulativa konstruktioner, utan utgick från sina egna observationer och förlitade sig på de verkliga egenskaperna hos levande organismer. Samtidigt berikade hon de biologiska verktygen med den historiska metoden.
Formuleringen av evolutionsteorin orsakade inte bara en stormig vetenskaplig diskussion, utan gav också impulser till utvecklingen av sådana vetenskaper som allmän biologi, genetik, avel, antropologi och en rad andra. I detta avseende kan man inte annat än instämma i påståendet att evolutionsteorin krönte nästa steg i biologins utveckling och blev startpunkten för dess framsteg under 1900-talet.
Bevis för utvecklingen av vilda djur. Evolutionära resultat: organismers anpassningsförmåga till sin miljö, mångfald av arter
Bevis för utvecklingen av vilda djur
Inom olika biologiområden, även före Charles Darwin och efter publiceringen av hans evolutionsteori, erhölls ett antal bevis för att stödja den. Dessa vittnesmål kallas bevis på evolution... De oftast citerade är paleontologiska, biogeografiska, jämförande embryologiska, jämförande anatomiska och jämförande biokemiska bevis på evolution, även om taxonomi och växt- och djuruppfödningsdata inte kan bortses från.
Paleontologiska bevis baserat på studier av fossila rester av organismer. Dessa inkluderar inte bara välbevarade organismer frusna i is eller inneslutna i bärnsten, utan också "mumier" som finns i sura torvmossar, samt rester av organismer och fossiler som bevaras i sedimentära bergarter. Förekomsten av enklare organismer i gamla bergarter än i senare lager, och det faktum att arter som finns på en nivå försvinner på en annan, anses vara ett av de viktigaste bevisen på evolution och förklaras av arternas uppkomst och utrotning under motsvarande epoker p.g.a. förändringar i miljöförhållandena.
Trots att det hittills har hittats få fossila lämningar och många fragment saknas i fossilregistret på grund av den låga sannolikheten för att bevara organiska lämningar, har man hittat former av organismer som har tecken på både evolutionärt äldre och yngre grupper av organismer. Sådana former av organismer kallas övergångsformer... Livliga representanter för övergångsformer, som illustrerar övergången från fisk till landlevande ryggradsdjur, är korsfenade fiskar och stegocephaler, och Archaeopteryx upptar en viss plats mellan reptiler och fåglar.
Serien av fossila former, som konsekvent är förbundna med varandra i evolutionsprocessen, inte bara genom allmänna utan också genom särskilda egenskaper hos strukturen, kallas fylogenetisk serie... De kan representeras av fossila lämningar från olika kontinenter, och hävdar att de är mer eller mindre kompletta, men deras studie är omöjlig utan jämförelse med levande former för att påvisa utvecklingen av evolutionsprocessen. Ett klassiskt exempel på en fylogenetisk serie är evolutionen av hästens förfäder, studerad av grundaren av evolutionär paleontologi V.O.Kovalevsky.
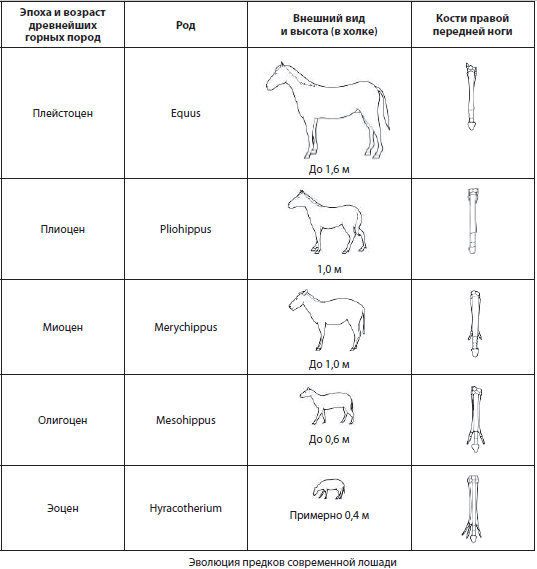
Biogeografiska bevis. Biogeografi hur vetenskapen studerar distributions- och distributionsmönstren över vår planets yta av arter, släkten och andra grupper av levande organismer, såväl som deras samhällen.
Frånvaron i någon del av jordens yta av arter av organismer som är anpassade till ett sådant habitat och som slår väl rot med artificiell import, som kaniner i Australien, samt närvaron av nära former av organismer i delar av landet som är åtskilda kl. avsevärda avstånd från varandra indikerar först och främst att jordens utseende inte alltid var detsamma, och geologiska omvandlingar, i synnerhet kontinenternas drift, bildandet av berg, uppgång och fall av världens nivå Ocean, påverka utvecklingen av organismer. Till exempel, i de tropiska regionerna i Sydamerika, Sydafrika och Australien finns det fyra liknande arter av lungfisk, medan områdena med kameler och lamor som tillhör samma ordning finns i Nordafrika, större delen av Asien och Sydamerika. Paleontologiska studier har visat att kameler och lamor härstammar från en gemensam förfader som en gång levde i Nordamerika och spred sig sedan till Asien genom den redan existerande näset vid platsen för Beringssundet, såväl som över Panamanäset till Sydamerika . Därefter dog alla representanter för denna familj ut i mellanområdena, och i marginalområdena bildades nya arter i evolutionsprocessen. Den tidigare separationen av Australien från resten av landmassorna gjorde det möjligt att där bilda en helt speciell flora och fauna, i vilken sådana former av däggdjur som monotremes - näbbdjuret och echidna - har överlevt.
Ur biogeografins synvinkel är det möjligt att förklara mångfalden av Darwins finkar på Galapagosöarna, 1200 km från Sydamerikas kust och med vulkaniskt ursprung. Tydligen flög representanter för den enda arten av finkar i Ecuador en gång på dem eller introducerades, och sedan, när de förökade sig, bosatte sig några av individerna på resten av öarna. På centralen stora öar kampen för tillvaron (mat, häckningsplatser, etc.) var den mest akuta, därför bildades arter som skilde sig något från varandra i utseende, som konsumerade olika livsmedel (frön, frukter, nektar, insekter, etc.).
Påverkade spridningen av olika grupper av organismer och förändringar i klimatförhållandena på jorden, vilket bidrog till vissa gruppers välstånd och andras utrotning. Enskilda arter eller grupper av organismer som har överlevt från tidigare utbredd flora och fauna kallas reliker... Dessa inkluderar ginkgo, sequoia, tulpanträd, korsfenad coelacanth fisk, etc. I en vidare bemärkelse de arter av växter och djur som lever på begränsade områden territorier eller vatten kallas endemisk, eller endemisk... Till exempel är alla representanter för den inhemska floran och faunan i Australien endemiska, och i Bajkalsjöns flora och fauna finns det upp till 75% av dem.
Jämförande anatomiska bevis. Studiet av anatomin hos besläktade grupper av djur och växter ger övertygande bevis på likheten i strukturen hos deras organ. Trots det faktum att miljöfaktorer verkligen sätter sin prägel på organens struktur, hos angiospermer, med all sin fantastiska mångfald, har blommor foderblad, kronblad, ståndare och pistiller, och hos landlevande ryggradsdjur är lemmen byggd enligt en femtåig plan . Organ som har en liknande struktur, intar samma position i kroppen och utvecklas från samma primordia i besläktade organismer, men utför olika funktioner, kallas homolog... Sålunda är hörselbenen (malleus, incus och stapes) homologa med fiskens grenbågar, giftkörtlarna hos ormar liknar spottkörtlarna hos andra ryggradsdjur, däggdjursbröstkörtlar är svettkörtlar, sälar och valar. är homologa med fåglars vingar, lemmar på hästar och mullvadar.

De organ som inte fungerar under lång tid, troligen, i evolutionsprocessen förvandlas till rudimentär (rudiment)- strukturer underutvecklade i jämförelse med förfäders former, som har förlorat sin huvudsakliga betydelse. Dessa inkluderar skenbenet hos fåglar, ögon hos mullvadar och mullvadsråttor, hår, svanskotan och blindtarm hos människor, etc.
Hos vissa individer kan det dock finnas tecken som saknas hos denna art, men som fanns hos avlägsna förfäder - atavismer till exempel tretålighet hos moderna hästar, utveckling av ytterligare par bröstkörtlar, svans och hårfäste i hela människokroppen.
Om homologa organ är bevis för släktskap mellan organismer och divergens i evolutionsprocessen, då liknande kroppar- strukturer som liknar organismer av olika grupper, utför samma funktioner, tvärtom, hänvisa till exempel konvergens(konvergens kallas i allmänhet den oberoende utvecklingen av liknande egenskaper i olika grupper av organismer som existerar under samma förhållanden) och bekräftar det faktum att miljön lämnar ett betydande avtryck på organismen. Analoger är insekters och fåglars vingar, ögonen på ryggradsdjur och bläckfiskar (bläckfiskar, bläckfiskar), ledade lemmar hos leddjur och landlevande ryggradsdjur.

Jämförande embryologiska bevis. Genom att studera embryonal utveckling hos representanter för olika grupper av ryggradsdjur upptäckte K. Baer deras slående strukturella enhet, särskilt i de tidiga utvecklingsstadierna ( embryonal likhetslag). Senare formulerade E. Haeckel biogenetisk lag, enligt vilken ontogeni är en kort upprepning av fylogenesen, det vill säga de stadier som en organism går igenom i processen för sin individuella utveckling upprepar den historiska utvecklingen av den grupp som den tillhör.
Således får embryot hos ett ryggradsdjur i de första utvecklingsstadierna strukturella egenskaper som är karakteristiska för fiskar, och sedan för groddjur och i slutändan för den grupp som den tillhör. Denna omvandling förklaras av det faktum att var och en av ovanstående klasser har gemensamma förfäder med moderna reptiler, fåglar och däggdjur.

Den biogenetiska lagen har dock ett antal begränsningar, och därför begränsade den ryske forskaren A. N. Severtsov avsevärt tillämpningsområdet för dess tillämpning genom att uteslutande upprepa funktionerna i de embryonala utvecklingsstadierna av förfäders former i ontogenes.
Jämförande biokemiska bevis. Utvecklingen av mer exakta metoder för biokemisk analys försåg evolutionära forskare med en ny grupp av data till förmån för den historiska utvecklingen av den organiska världen, eftersom närvaron av samma ämnen i alla organismer indikerar en möjlig biokemisk homologi, liknande den vid nivå av organ och vävnader. Jämförande biokemiska studier av den primära strukturen hos sådana utbredda proteiner som cytokrom med och hemoglobin, såväl som nukleinsyror, särskilt rRNA, visade att många av dem har praktiskt taget samma struktur och utför samma funktioner hos representanter för olika arter, medan ju närmare förhållandet är, desto större likhet finns i strukturen av studerade ämnen.
Således bekräftas evolutionsteorin av en betydande mängd data från olika källor, vilket återigen vittnar om dess tillförlitlighet, men den kommer fortfarande att förändras och förfinas, eftersom många aspekter av organismernas liv förblir utanför synfältet för forskare.
Evolutionära resultat: organismers anpassningsförmåga till sin miljö, mångfald av arter
Förutom de allmänna egenskaperna som är karakteristiska för representanter för ett visst rike, kännetecknas typerna av levande organismer av en fantastisk variation av funktioner i den yttre och inre strukturen, vital aktivitet och till och med beteende, som dök upp och valdes ut i evolutionsprocessen och säkerställa anpassning till livsmiljöförhållanden. Det bör dock inte antas att eftersom fåglar och insekter har vingar så beror detta på luftmiljöns direkta inverkan, eftersom det finns gott om vinglösa insekter och fåglar. De tidigare nämnda anpassningarna valdes genom naturligt urval från hela spektrumet av tillgängliga mutationer.
Epifytiska växter, som inte lever på jord, utan på träd, har anpassat sig till absorptionen av atmosfärisk fukt med rötter utan rothår, men med en speciell hygroskopisk vävnad - velamen... Vissa bromelia kan absorbera vattenånga i den fuktiga atmosfären i tropikerna med hjälp av hår på bladen.
Insektsätande växter (soldagg, Venus flugfälla), som lever på jordar där kväve inte är tillgängligt av en eller annan anledning, har utvecklat en mekanism för att attrahera och absorbera små djur, oftast insekter, som är källan till det önskade elementet för dem.
För att skydda mot att äta av växtätare har många växter som leder ett fast liv utvecklat passiva försvarsmedel, såsom taggar (hagtorn), taggar (ros), stickande hårstrån (nässlor), ansamling av kristaller av kalciumoxalat (sorrel), biologiskt aktiva ämnen i vävnader (kaffe, hagtorn) etc. I vissa av dem är till och med frön i omogna frukter omgivna av steniga celler som hindrar skadedjur från att nå dem, och först till hösten sker lignifieringsprocessen, vilket gör att fröna tillåter att komma ner i jorden och gro (päron).
Miljön har en bildande effekt även på djur. Så många fiskar och vattenlevande däggdjur har en strömlinjeformad kropp, vilket gör det lättare för dem att röra sig i sin tjocklek. Man ska dock inte anta att vatten direkt påverkar kroppens form, bara i evolutionsprocessen var det just de djuren som hade denna egenskap som var mest anpassade till det.
Samtidigt är kroppen av valar och delfiner inte täckt med hår, medan den besläktade gruppen av pinnipeds har en något reducerad päls, eftersom de, till skillnad från de förra, tillbringar en del av tiden på land, där utan ull skulle deras hud blir genast isig...
Kroppen på de flesta fiskar är täckt med fjäll, som är ljusare på undersidan än på den övre, vilket gör att dessa djur knappast är märkbara ovanifrån för naturliga fiender mot bakgrund av botten och underifrån - mot botten. bakgrund av himlen. Färgning som säkerställer osynligheten av djur för deras fiender eller offer kallas nedlåtande... Den är utbredd i naturen. Ett slående exempel på en sådan färgning är färgningen av undersidan av vingarna på en callima-fjäril, som, sittande på en kvist och viker ihop vingarna, visar sig vara som ett torrt löv. Andra insekter, som stickinsekter, förkläder sig till växtkvistar.
Fläckig eller randig färg är också anpassningsbar, eftersom fåglar som vaktlar eller ejder inte kan ses mot bakgrund av jorden, ens på nära håll. Prickiga ägg från fåglar som häckar på marken är också osynliga.
Djurens färg är inte alltid lika konstant som för en zebra, till exempel kan en flundra och en kameleont ändra den beroende på arten av den plats där de befinner sig. Gökar, som lägger sina ägg i olika fåglars bon, kan variera färgen på sina skal så att boets "ägare" inte märker skillnaderna mellan dem och sina egna ägg.
Färgen på djur kan inte alltid göra dem osynliga - många av dem är helt enkelt slående, vilket borde varna för fara. De flesta av dessa insekter och reptiler är giftiga i en eller annan grad, som till exempel en nyckelpiga eller en geting, därför ett rovdjur, som har upplevt flera gånger obehag efter att ha ätit ett sådant föremål, undviker det. Ändå, varningsfärgningär inte universell, eftersom vissa fåglar har anpassat sig för att livnära sig på dem (getingätare).
De ökade chanserna att överleva hos individer med varnande färgning bidrog till att den uppträdde hos representanter för andra arter utan vederbörlig anledning. Detta fenomen kallas härmning... Så, icke-giftiga larver av vissa arter av fjärilar imiterar giftiga, och nyckelpigor- en av typerna av kackerlackor. Däremot kan fåglar snabbt lära sig att skilja giftiga från icke-giftiga organismer och konsumera de sistnämnda, och undvika individer som fungerade som en förebild.
I vissa fall kan det motsatta fenomenet också observeras - rovdjur imiterar ofarliga i färg, vilket gör att de kan närma sig offret på nära avstånd och sedan attackera (sabeltandad blandning).
Skydd för många arter tillhandahålls också av adaptivt beteende, som är förknippat med att lagra mat för vintern, ta hand om avkommor, frysa på plats eller vice versa, anta en hotfull pose. Till exempel förbereder flodbävrar för vintern flera kubikmeter grenar, delar av stammar och annan växtmat, och svämmar över det i vattnet nära "hydorna".
Att ta hand om avkomma är huvudsakligen inneboende hos däggdjur och fåglar, men det finns också i representanter för andra klasser av chordater. Till exempel är klibbalhanarnas aggressiva beteende känt, som driver bort alla fiender från boet där äggen finns. Manliga grodor lindar ägg på tassarna och bär dem tills grodyngeln kläcks från dem.
Även vissa insekter kan ge sin avkomma en mer gynnsam livsmiljö. Till exempel matar bin sina larver, medan unga bin till en början bara "arbetar" i kupan. Myror flyttar sina puppor upp och ner i boet, beroende på temperatur och luftfuktighet, och när det finns risk för översvämning tar de dem vanligtvis med sig. Skalbaggar förbereder speciella bollar för sina larver från djuravfall.
När de hotas av attack fryser många insekter på plats och tar formen av torra pinnar, kvistar och löv. Och huggormar, tvärtom, reser sig och blåser upp sin huva, medan en skallerorm gör ett speciellt ljud med en skallra i slutet av svansen.
Beteendeanpassningar kompletteras med fysiologiska anpassningar förknippade med miljöns egenskaper. Så en person kan vara under vatten utan dykutrustning i bara några minuter, varefter han kan förlora medvetandet och dö på grund av syrebrist, och valar dyker inte upp under tillräckligt lång tid. Deras lungkapacitet är inte för stor, men det finns andra fysiologiska anpassningar, till exempel i musklerna finns en hög koncentration av andningspigment - myoglobin, som så att säga lagrar syre och ger bort det under dykning. Dessutom har valar en speciell formation - ett "mirakulöst nätverk" som tillåter användning av syre även från venöst blod.
Djur i varma livsmiljöer, som öknar, riskerar ständigt att överhettas och förlora överflödig fukt. Därför har fennec räven extremt stora öronen som gör att den kan utstråla värme. För att undvika fuktförlust genom huden, tvingas amfibier i ökenregioner att byta till en nattlig livsstil när luftfuktigheten stiger och dagg dyker upp.
Fåglar som har bemästrat lufthabitatet, förutom anatomiska och morfologiska anpassningar för flygning, har också viktiga fysiologiska egenskaper. Till exempel, på grund av det faktum att rörelse i luften kräver en extremt stor energiförbrukning, kännetecknas denna grupp av ryggradsdjur av en hög ämnesomsättning, och de utsöndrade ämnesomsättningsprodukterna utsöndras omedelbart, vilket bidrar till en minskning av den specifika densiteten av kroppen.
Anpassningar till miljön, trots all perfektion, är relativa. Så, vissa arter av milkweed producerar alkaloider som är giftiga för de flesta djur, men larverna från en av arterna av fjärilar - danaiderna - livnär sig inte bara på milkweed-vävnader, utan ackumulerar också dessa alkaloider och blir oätliga för fåglar.
Dessutom är anpassningar endast tillrådliga i en viss livsmiljö och är värdelösa i en annan miljö. Till exempel sällsynta och stort rovdjur Ussuri-tigern, som alla katter, har mjuka dynor på tassarna och indragbara vassa klor, vassa tänder, utmärkt syn även i mörker, skarp hörsel och starka muskler, vilket gör att den kan upptäcka sitt byte, smyga sig på det och lägga ett bakhåll för det. . Dess randiga färg döljer den dock bara på våren, sommaren och hösten, medan den i snön blir tydligt synlig och tigern bara kan räkna med en blixtattack.
Fikonblomställningar, som ger värdefull infructescence, har en så specifik struktur att de endast pollineras av getingar med blastfager, och därför, introducerade i kulturen, bar de inte frukt under lång tid. Endast förädling av partenokarpiska varianter av fikon (bildar frukter utan befruktning) kunde rädda situationen.
Trots att exempel på artbildning beskrivs under mycket korta tidsperioder, som i fallet med skallran på de kaukasiska ängarna, som på grund av regelbunden slåtter först delades upp i två populationer - tidig blomning och fruktsättning och sen blomning, i Faktum är att mikroevolutionen med största sannolikhet kräver mycket längre perioder - många århundraden, eftersom mänskligheten, vars olika grupper har slitits bort från varandra i tusentals år, ändå inte har delats upp i olika arter. Men eftersom evolutionen har praktiskt taget obegränsad tid, i hundratals miljoner och miljarder år, har flera miljarder arter redan levt på jorden, varav de flesta har dött ut, och de som har kommit ner till oss är kvalitativa stadier av denna kontinuerliga process.
Enligt moderna data finns det över 2 miljoner arter av levande organismer på jorden, varav de flesta (cirka 1,5 miljoner arter) tillhör djurriket, cirka 400 tusen till växtriket, över 100 tusen till svampriket, och vila - till bakterier. Denna slående mångfald är resultatet av arternas divergens (divergens) för olika morfologiska, fysiologiska, biokemiska, ekologiska, genetiska och reproduktiva egenskaper. Till exempel inkluderar en av de största växtsläktena, som tillhör orkidéfamiljen, - dendrobium - över 1 400 arter och släktet kaloidbaggar - över 1 600 arter.
Klassificeringen av organismer är systematikens uppgift, som i 2 tusen år har försökt bygga inte bara en harmonisk hierarki, utan ett "naturligt" system som återspeglar graden av släktskap mellan organismer. Emellertid har alla försök att göra detta ännu inte krönts med framgång, eftersom i ett antal fall, under evolutionens gång, inte bara skillnader mellan karaktärer observerades, utan också konvergens (konvergens), som ett resultat av vilket, i mycket avlägsna grupper, organ förvärvade egenskaper av likhet, såsom till exempel ögonen på bläckfiskar och däggdjursögon.
Makrovolution. Riktningar och evolutionsmetoder (A. N. Severtsov, I. I. Shmalgauzen). Biologiska framsteg och regression, aromorfos, idioadaptation, degeneration. Orsaker till biologiska framsteg och regression. Hypoteser om livets ursprung på jorden. De viktigaste aromorfoserna i utvecklingen av växter och djur. Komplikation av levande organismer i evolutionsprocessen
Makrovolution
Bildandet av en art markerar en ny omgång av den evolutionära processen, eftersom individer av denna art, som är mer anpassade till miljöförhållanden än individer av moderarten, gradvis bosätter sig i nya territorier och redan i dess populationer mutagenes, befolkningsvågor, isolering och naturligt urval spelar sin kreativa roll. ... Med tiden ger dessa populationer upphov till nya arter, som på grund av genetisk isolering har mycket mer tecken på likhet med varandra än med arterna av det släkte som artens förfader spunnet av, och därmed uppstår ett nytt släkte, då en ny familj, ordning (ordning), klass, etc. Uppsättningen av evolutionära processer som leder till uppkomsten av supraspecifika taxa (släkten, familjer, ordnar, klasser, etc.) kallas makroevolution. Makroevolutionära processer generaliserar så att säga mikroevolutionära förändringar som inträffar under lång tid, samtidigt som de avslöjar de viktigaste trenderna, riktningarna och mönstren för utvecklingen av den organiska världen, som inte kan observeras på en lägre nivå. Hittills har inga specifika mekanismer för makroevolution identifierats, därför tror man att den endast utförs genom mikroevolutionära processer, men denna position utsätts ständigt för välgrundad kritik.
Framväxten av ett komplext hierarkiskt system i den organiska världen är till stor del resultatet av den ojämna evolutionen av olika grupper av organismer. Så, den redan nämnda ginkgo biloba, så att säga, "bevarad" i tusentals år, medan tallar ganska nära den har förändrats avsevärt under denna tid.
Riktningar och evolutionsmetoder (A. N. Severtsov, I. I. Shmalgauzen). Biologiska framsteg och regression, aromorfos, idioadaptation, degeneration
Genom att analysera den organiska världens historia kan man märka att med vissa intervaller rådde separata grupper av organismer, som sedan tenderade att avta eller helt försvinna. Således kan tre stamlinjer urskiljas. evolutionens riktningar: biologiska framsteg, biologisk regression och biologisk stabilisering. De ryska evolutionisterna A. N. Severtsov och I. I. Shmalgauzen gav ett betydande bidrag till utvecklingen av läran om evolutionens riktningar och vägar.
Biologiska framsteg förknippas med det biologiska välståndet för gruppen som helhet och kännetecknar dess evolutionära framgång. Den speglar den naturliga utvecklingen av levande natur från enkel till komplex, från en lägre grad av organisation till en högre. Enligt AN Severtsov är kriterierna för biologiska framsteg ökningen av antalet individer i en given grupp, utvidgningen av dess utbredningsområde, såväl som uppkomsten och utvecklingen av grupper av lägre rang inom den (omvandling av en art till en släkte, ett släkte in i en familj, etc.). För närvarande observeras biologiska framsteg hos angiospermer, insekter, teleostfiskar och däggdjur.
Enligt A. N. Severtsov kan biologiska framsteg uppnås som ett resultat av vissa morfofysiologiska transformationer av organismer, medan han identifierade tre huvudsakliga sätt att uppnå: arogenes, allogenes och katagenes.

Arogenes, eller morfofysiologiska framsteg, är förknippad med en betydande utvidgning av utbudet av denna grupp av organismer på grund av förvärvet stora förändringar strukturer - aromorfoser.
Aromorfos kallas den evolutionära omvandlingen av en organisms struktur och funktioner, vilket ökar dess organisationsnivå och öppnar för nya möjligheter för anpassning till olika existensvillkor.
Exempel på aromorfoser är uppkomsten av en eukaryot cell, multicellularitet, uppkomsten av ett hjärta hos fiskar och dess delning med en fullständig septum hos fåglar och däggdjur, bildandet av en blomma i angiospermer, etc.
Allogenes, till skillnad från arogenes, åtföljs inte av en utvidgning av området, men inom det gamla finns en betydande variation av former som har särskilda anpassningar till miljön - idioadaptation.
Idioanpassning– Det här är en liten morfofysiologisk anpassning till speciella miljöförhållanden, användbar i kampen för tillvaron, men förändrar inte organisationsnivån. Dessa förändringar illustreras av skyddande färg hos djur, en mängd olika mundelar hos insekter, taggar av växter, etc. Inte mindre ett bra exempelär Darwins finkar, specialiserade på olika typer av mat, där transformationer först påverkade näbben och sedan andra delar av kroppen - fjäderdräkt, svans, etc.
Paradoxalt nog kan förenklingen av organisationen leda till biologiska framsteg. Denna väg kallas katagenes.
Degenerationär en förenkling av organismer i evolutionsprocessen, som åtföljs av förlusten av vissa funktioner eller organ.
Den biologiska utvecklingens fas ersätts av en fas biologisk stabilisering, vars essens är att bevara egenskaperna hos en given art som den mest gynnsamma i en given mikromiljö. Enligt II Shmalgauzen betyder det inte alls att evolutionen upphör, tvärtom betyder det organismens maximala överensstämmelse med förändringar i miljön. De "levande fossilerna" av coelacanth, gingko och andra är i fasen av biologisk stabilisering.
Antipoden för biologiska framsteg är biologisk regression- den evolutionära nedgången för denna grupp på grund av oförmågan att anpassa sig till miljöförändringar. Det visar sig i en minskning av antalet befolkningar, en minskning av områden, en minskning av antalet grupper av lägre rang i sammansättningen högre taxon... En grupp organismer som befinner sig i ett tillstånd av biologisk regression är hotad av utrotning. Många exempel på detta fenomen kan ses i den organiska världens historia, och för närvarande är regression karakteristisk för vissa ormbunkar, amfibier och reptiler. Med människans tillkomst orsakas biologisk regression ofta av hennes ekonomiska aktiviteter.
Den organiska världens utvecklingsriktningar och -vägar utesluter inte varandra, det vill säga utseendet av aromorfos betyder inte att idioadaptation eller degeneration inte längre kan inträffa. Tvärtom, enligt den utvecklade av A.N.Severtsov och I.I.Shmalgauzen fasändringsregel, olika riktningar av den evolutionära processen och sätt att uppnå biologiska framsteg ersätter naturligt varandra. Under evolutionens gång kombineras dessa vägar: ganska sällsynta aromorfoser överför en grupp organismer till en kvalitativt ny organisationsnivå, och i framtiden följer den historiska utvecklingen vägen för idioadaptation eller degeneration, vilket säkerställer anpassning till specifika miljöförhållanden.
Orsaker till biologiska framsteg och regression
I evolutionsprocessen övervinns gränsen för naturligt urval och följaktligen framskrider endast de grupper av organismer där ärftlig variation skapar ett tillräckligt antal kombinationer som kan säkerställa överlevnaden för gruppen som helhet.
Samma grupper som av någon anledning inte har ett sådant reservat är i de flesta fall dömda till utrotning. Detta beror ofta på lågt urvalstryck i de tidigare stadierna av evolutionsprocessen, vilket ledde till en snäv specialisering av gruppen eller till och med degenerativa fenomen. Konsekvensen av detta är omöjligheten att anpassa sig till nya miljöförhållanden under dess plötsliga förändringar. Ett slående exempel på detta är dinosauriernas plötsliga död på grund av en gigantisk himlakropps fall till jorden för 65 miljoner år sedan, vilket resulterade i en jordbävning, höjning av miljontals ton damm i luften, en kraftig nedkylning, döden för de flesta växter och växtätande djur. Samtidigt kunde de moderna däggdjurens förfäder, som inte hade några snäva preferenser för matkällor och var varmblodiga, överleva dessa förhållanden och inta en dominerande ställning på planeten.
Hypoteser om livets ursprung på jorden
Av hela spektrumet av hypoteser för bildandet av jorden vittnar det största antalet fakta till förmån för teorin om "Big Bang". Med tanke på det faktum att detta vetenskapliga antagande huvudsakligen bygger på teoretiska beräkningar, har Large Hadron Collider, byggd i European Center kärnkraftsforskning nära Genève (Schweiz). Enligt Big Bang-teorin bildades jorden för över 4,5 miljarder år sedan tillsammans med solen och andra planeter i solsystemet som ett resultat av kondensation av ett gas- och dammmoln. En minskning av planetens temperatur och migrationen av kemiska element på den bidrog till dess stratifiering i kärnan, manteln och skorpan, och de efterföljande geologiska processerna (rörelse kontinentalplattor, vulkanisk aktivitet, etc.) orsakade bildandet av atmosfären och hydrosfären.
Liv har också funnits på jorden under mycket lång tid, vilket framgår av fossila rester av olika organismer i stenar, men fysikaliska teorier kan inte svara på frågan om tidpunkten och orsakerna till dess förekomst. Det finns två motsatta synpunkter på livets ursprung på jorden: teorin om abiogenes och biogenes. Abiogenes teorier hävda möjligheten av de levandes ursprung från de icke-levande. Dessa inkluderar kreationism, hypotesen om spontan generering och teorin om biokemisk evolution av A.I. Oparin.
Grundläggande ställning kreationism var världens skapelse av någon övernaturlig varelse (Skapare), vilket återspeglas i myterna om världens folk och religiösa kulter, men planetens ålder och livet på den är mycket högre än de datum som anges i dessa källor , och det finns massor av inkonsekvenser i dem.
Grundaren spontan generation teori livet anses vara den antika grekiska vetenskapsmannen Aristoteles, som hävdade att flera uppkomsten av nya varelser är möjlig, till exempel daggmaskar från pölar och maskar och flugor från ruttet kött. Dessa åsikter motbevisades emellertid under 1600- och 1800-talen av F. Redis och L. Pasteurs modiga experiment.
Den italienska läkaren Francesco Redi placerade 1688 köttbitar i krukor och förseglade dem tätt, men inga maskar hölls i dem, medan de dök upp i öppna krukor. För att motbevisa den då rådande uppfattningen att livsprincipen finns i luften, upprepade han sina experiment, men förseglade inte krukorna, utan stängde dem med flera lager muslin, och livet dök inte upp igen. Trots de övertygande data som F. Redi erhållit, gav studierna av A. van Leeuwenhoek ny mat för diskussioner om "livets början", som fortsatte under nästa århundrade.
En annan italiensk forskare, Lazzaro Spallanzani, modifierade F. Redis experiment 1765 genom att koka kött och grönsaksavkok i flera timmar och försegla dem. Efter flera dagar hittade han inte heller några tecken på liv där och drog slutsatsen att det levande bara kan uppstå från det levande.
Det sista slaget mot teorin om spontan spontan generation slogs av den store franske mikrobiologen Louis Pasteur 1860, som placerade kokt buljong i en kolv med en S-formad hals och inte fick några embryon. Det verkar som om detta vittnade till förmån för teorierna om biogenes, men frågan förblev öppen om hur den allra första organismen uppstod.
Den sovjetiska biokemisten A.I. Oparin försökte svara på det, som kom till slutsatsen att sammansättningen av jordens atmosfär i de tidiga stadierna av dess existens inte alls var densamma som i vår tid. Troligtvis bestod den av ammoniak, metan, koldioxid och vattenånga, men innehöll inte fritt syre. Under påverkan av elektriska urladdningar med hög effekt och vid höga temperaturer, det enklaste organiska föreningar, vilket bekräftades av experimenten av S. Miller och G. Urey 1953, som erhöll från de ovan nämnda föreningarna flera aminosyror, enkla kolhydrater, adenin, urea, samt de enklaste fettsyrorna, myr- och ättiksyrorna.
Ändå betyder syntesen av organiska ämnen ännu inte uppkomsten av liv, därför lade A.I. Oparin fram hypotes om biokemisk evolution, enligt vilka olika organiska ämnen uppstod och förenades till större molekyler i havens och oceanernas grunda vatten, där förutsättningarna för kemisk syntes och polymerisation är mest gynnsamma. RNA-molekyler anses nu vara de första bärarna av liv.
Vissa av dessa ämnen bildade gradvis stabila komplex i vatten - koacervater, eller koacervata droppar som liknar fettdroppar i buljong. Dessa koacervat fick en mängd olika ämnen från den omgivande lösningen, som genomgick kemiska omvandlingar i dropparna. Liksom organiska ämnen var koacervat i sig inte levande varelser, utan var nästa steg i deras uppkomst.
De av koacervaten som hade ett bra förhållande av ämnen i sin sammansättning, särskilt proteiner och nukleinsyror, på grund av enzymproteinernas katalytiska egenskaper, fick med tiden förmågan att reproducera sin egen typ och utföra metaboliska reaktioner, samtidigt som strukturen hos proteiner kodades av nukleinsyror.
Men, förutom reproduktion, kännetecknas levande system av beroende av tillförsel av energi utifrån. Detta problem löstes till en början på grund av den syrefria nedbrytningen av organiska ämnen från miljön (det fanns inget syre i atmosfären vid den tiden), d.v.s.
heterotrofisk näring. En del av de absorberade organiska ämnena kunde samla energin från solljus, som klorofyll, vilket gjorde det möjligt för ett antal organismer att gå över till autotrof näring. Utsläppet av syre i atmosfären under fotosyntesen ledde till uppkomsten av effektivare syreandning, uppkomsten av ozonskiktet och, i slutändan, utsläppet av organismer till land.
Följaktligen var resultatet av den kemiska utvecklingen uppkomsten protobioter- primära levande organismer, från vilka alla för närvarande existerande arter har sitt ursprung som ett resultat av biologisk evolution.
Teorin om biokemisk evolution i vår tid är den mest bekräftade, men idén om de specifika mekanismerna för livets ursprung har förändrats. Till exempel visade det sig att bildandet av organiska ämnen börjar i rymden, och organiska ämnen spelar en viktig roll även i själva bildandet av planeter, vilket säkerställer vidhäftning av små delar. Dessutom sker bildandet av organiskt material i planetens tarmar: med ett utbrott skjuter vulkanen ut upp till 15 ton organiskt material. Det finns andra hypoteser om mekanismerna för koncentration av organiska ämnen: frysning av en lösning, absorption (bindning) på ytan av vissa mineralföreningar, verkan av naturliga katalysatorer, etc. Uppkomsten av liv på jorden är för närvarande omöjlig, eftersom allt organiskt material som bildas spontant vid någon punkt skulle planeter omedelbart oxideras av atmosfärens fria syre eller användas av heterotrofa organismer. Detta förstod redan 1871 av Charles Darwin.
Biogenes teorier förneka livets spontana ursprung. De viktigaste är steady state-hypotesen och panspermihypotesen. Den första av dem är baserad på det faktum att livet existerar för evigt, ändå finns det mycket gamla stenar på vår planet, där det inte finns några spår av aktiviteten i den organiska världen.
Panspermihypotes hävdar att livets embryon fördes till jorden från rymden av några utomjordingar eller av gudomlig försyn. Denna hypotes stöds av två fakta: behovet av allt levande, vilket är ganska sällsynt på planeten, men som ofta finns i meteoriter, molybden, samt upptäckten av organismer som liknar bakterier på meteoriter från Mars. Hur livet uppstod på andra planeter är dock oklart.
De viktigaste aromorfoserna i utvecklingen av växter och djur
Växt- och djurorganismer, som representerar olika grenar av utvecklingen av den organiska världen, förvärvade självständigt vissa strukturella egenskaper i processen för historisk utveckling, som kommer att karakteriseras nedan.
Hos växter är de viktigaste av dem övergången från haploidi till diploidi, oberoende från vatten under befruktningsprocessen, övergången från extern till inre befruktning och uppkomsten av dubbel befruktning, sönderdelning av kroppen i organ, utveckling av ledningssystemet, komplikation och förbättring av vävnader, samt specialisering av pollinering med hjälp av insekter och spridning av frön och frukter.
Övergången från haploidi till diploidi gjorde växter mer motståndskraftiga mot miljöfaktorer på grund av en minskning av risken för recessiva mutationer. Tydligen påverkade denna transformation förfäderna till kärlväxter, som inte inkluderar mossor, som kännetecknas av övervikten av gametofyten i livscykeln.
De viktigaste aromorfoserna i djurens evolution är förknippade med uppkomsten av multicellularitet och den ökande sönderdelningen av alla organsystem, uppkomsten av ett starkt skelett, utvecklingen av det centrala nervsystemet, såväl som socialt beteende hos olika grupper av högorganiserade djur, som gav impulser till mänskliga framsteg.
Komplikation av levande organismer i evolutionsprocessen
Historien om den organiska världen på jorden studeras av de överlevande kvarlevorna, avtrycken och andra spår av levande organismers vitala aktivitet. Hon är ämnet vetenskap paleontologi... Baserat på det faktum att resterna av olika organismer finns i olika lager av bergarter skapades en geokronologisk skala, enligt vilken jordens historia delades in i vissa tidsperioder: eoner, epoker, perioder och århundraden.
Eon kallas en stor tidsperiod i geologisk historia, som förenar flera epoker. För närvarande särskiljs endast två eoner: kryptos (latent liv) och fanerozoikum (explicit liv). Epok– Det här är en tidsperiod i geologisk historia, som är en underavdelning av eonen, som i sin tur förenar perioder. I kryptozoikum särskiljs två epoker (arkeiska och proterozoikum), medan i fanerozoikum tre (paleozoikum, mesozoikum och kenozoikum).
En viktig roll i skapandet av den geokronologiska skalan spelades av ledande fossiler- resterna av organismer som var många med vissa intervall och välbevarade.
Utveckling av liv i kryptozoikum. Archaea och Proterozoic utgör det mesta av livets historia (period för 4,6 miljarder år - 0,6 miljarder år sedan), men det finns inte tillräckligt med information om livet under den perioden. De första resterna av organiskt material av biogent ursprung är cirka 3,8 miljarder år gamla, och prokaryota organismer fanns redan för 3,5 miljarder år sedan. De första prokaryoterna var en del av specifika ekosystem - cyanobakteriella mattor, på grund av vilken aktivitet specifika sedimentära bergarter av stromatoliter ("stenmattor") bildades.
Upptäckten av deras moderna analoger - stromatoliter i Shark Bay i Australien och specifika filmer på markytan i Sivash Bay i Ukraina - hjälpte till att förstå livet för långvariga prokaryota ekosystem. Fotosyntetiska cyanobakterier finns på ytan av cyanobakteriella mattor, och under deras lager finns extremt olika bakterier från andra grupper och arkéer. Mineraler som lägger sig på mattans yta och som bildas på grund av dess vitala aktivitet avsätts i lager (cirka 0,3 mm per år). Sådana primitiva ekosystem kan bara existera på platser som är olämpliga för andra organismers liv, och båda de ovan nämnda livsmiljöerna kännetecknas faktiskt av extremt hög salthalt.
Många data tyder på att jorden till en början hade en förnybar atmosfär, vilket inkluderade: koldioxid, vattenånga, svaveloxid, samt kolmonoxid, väte, svavelväte, ammoniak, metan, etc. Jordens första organismer var anaeroba, men tack vare fotosyntesen av cyanobakterier släpptes fritt syre ut i miljön, vilket först snabbt bunden med reduktionsmedel i miljön , och först efter bindning av alla reduktionsmedel började mediet få oxiderande egenskaper. Denna övergång bevisas av avsättningen av oxiderade former av järn - hematit och magnetit.
För cirka 2 miljarder år sedan, som ett resultat av geofysiska processer, flyttade nästan allt järn obundet i sedimentära bergarter till planetens kärna, och syre började ackumuleras i atmosfären på grund av frånvaron av detta element - en "syrerevolution" tog plats. Det var en vändpunkt i jordens historia, som inte bara innebar en förändring av atmosfärens sammansättning och bildandet av en ozonskärm i atmosfären - huvudförutsättningen för bosättning av land, utan också sammansättningen av bergarter bildas på jordens yta.
En annan viktig händelse ägde rum i Proterozoikum - uppkomsten av eukaryoter. På senare år har det varit möjligt att samla övertygande bevis för teorin om den eukaryota cellens endosymbiogenetiska ursprung – genom symbios av flera prokaryota celler. Förmodligen var den "huvudsakliga" förfadern till eukaryoter archaea, som bytte till absorption av matpartiklar genom fagocytos. Den ärftliga apparaten rörde sig in i cellens djup, samtidigt som förbindelsen med membranet bibehölls på grund av övergången av det uppkomna kärnhöljets yttre membran till membranen i det endoplasmatiska retikulumet.
Jordens geokronologiska historia Eon Epok Period Början för miljoner år sedan Varaktighet, miljoner år Livsutveckling Fanerozoisk kenozoisk antropogen 1,5 1,5 Fyra istid, följt av översvämningar, ledde till bildandet av köldbeständig flora och fauna (mammutar, myskoxar, ren lämlar). Utbytet av djur och växter mellan kontinenter på grund av uppkomsten av landbroar. Dominans av placenta däggdjur. Utrotning av många stora däggdjur. Bildandet av människan som biologisk art och dess spridning. Domesticering av djur och domesticering av växter. Försvinnande av många arter av levande organismer på grund av mänsklig ekonomisk aktivitet Neogen 25 23.5 Distribution av spannmål. Bildandet av alla moderna däggdjursordningar. Uppkomst stora apor Paleogen 65 40 Dominans av blommande växter, däggdjur och fåglar. Uppkomsten av klövdjur, köttätare, pinnipeds, primater, etc. Mesozoikum Mel 135 70 Uppkomsten av angiospermer, däggdjur och fåglar blir många Jura 195 60 Eran av reptiler och bläckfiskar. Uppkomsten av pungdjur och placenta däggdjur. Dominans av gymnospermer Trias 225 30 Första däggdjur och fåglar. Reptiler finns det gott om. Fördelning av örtartade sporer i den paleozoiska Perm 280 55 Framväxten av moderna insekter. Reptilutveckling. Utrotning av ett antal ryggradslösa grupper. Fördelning av barrträd Kol 345 65 De första reptilerna. Uppkomsten av bevingade insekter. Ormbunkar och åkerfräken råder.Devon 395 50 Fisk är rikligt. De första groddjuren. Uppkomsten av huvudgrupperna av sporer, första gymnospermer och svampar Silurian 430 35 Alger är riklig. De första landväxter och djur (spindlar). Vanliga käktåfiskar och kräftdjur Ordovicium 500 70 Koraller och trilobiter är rikligt förekommande. Blomning av gröna, bruna och röda alger. Bildandet av den första kordaten Kambrium 570 70 Många fiskfossiler. Sjöborrar och trilobiter är vanliga. Uppkomsten av flercelliga alger av Cryptose Proterose 2600 2000 Uppkomsten av eukaryoter. Främst encelliga grönalger är utbredda. Uppkomsten av multicellularitet. Utbrott av mångfald av flercelliga djur (uppkomsten av alla typer av ryggradslösa djur) Archaea 3500 1500 De första spåren av liv på jorden - bakterier och cyanobakterier. Uppkomsten av fotosyntes
Bakterierna som absorberades av cellen kunde inte smältas, utan förblev levande och fortsatte att fungera. Man tror att mitokondrier härstammar från lila bakterier som har förlorat förmågan att fotosyntetisera och som har oxiderat organiskt material. Symbios med andra fotosyntetiska celler har lett till bildandet av plastider i växtceller. Förmodligen uppstod flagellerna av eukaryota celler som ett resultat av symbios med bakterier, som, liksom moderna spiroketer, var kapabla att vrida rörelser. Till en början var den ärftliga apparaten hos eukaryota celler anordnad på ungefär samma sätt som i prokaryoter, och först senare, på grund av behovet av att kontrollera en stor och komplex cell, bildades kromosomer. Genomerna från intracellulära symbionter (mitokondrier, plastider och flageller) behöll i allmänhet sin prokaryota organisation, men de flesta av deras funktioner har gått över till kärngenomet.
Eukaryota celler har uppstått upprepade gånger och oberoende av varandra. Till exempel uppstod röda alger som ett resultat av symbiogenes med cyanobakterier och gröna alger - med proklorofytbakterier.
Resten av enkelmembranorganellerna och kärnan i den eukaryota cellen uppstod enligt endomembranteorin från invaginationerna av den prokaryota cellens membran.
Den exakta tiden för uppkomsten av eukaryoter är okänd, eftersom det redan finns avtryck av celler med liknande storlekar i cirka 3 miljarder år gamla avlagringar. På liknande sätt har eukaryoter registrerats i stenar som är cirka 1,5–2 miljarder år gamla, men först efter syrerevolutionen (för cirka 1 miljard år sedan) utvecklades gynnsamma förhållanden för dem.
I slutet av den proterozoiska eran (minst 1,5 miljarder år sedan) existerade redan flercelliga eukaryota organismer. Multicellularitet, som en eukaryot cell, har upprepade gånger förekommit i olika grupper av organismer.
Existerar olika synsätt om ursprunget för flercelliga djur. Enligt vissa data var deras förfäder flerkärniga, ciliatliknande celler, som sedan sönderföll till separata mononukleära celler.
Andra hypoteser kopplar ursprunget till flercelliga djur med differentieringen av koloniala encelliga celler. Avvikelserna mellan dem relaterar till utseendet av cellskikt i det ursprungliga flercelliga djuret. Enligt E. Haeckels hypotes om gastrea sker detta genom invaginering av en av väggarna hos en flercellig organism med ett lager, som i coelenterater. I motsats härtill formulerade I.I.Mechnikov fagocytellahypotesen, och ansåg enskikts sfäriska kolonier som volvox vara förfäder till flercelliga organismer, som absorberade matpartiklar genom fagocytos. Cellen som fångade partikeln förlorade sitt flagellum och flyttade djupt in i kroppen, där den genomförde matsmältningen, och i slutet av processen återvände den till ytan. Med tiden skedde en uppdelning av celler i två lager med vissa funktioner - den yttre gav rörelse och den inre - fagocytos. I. I. Mechnikov kallade en sådan organism fagocytella.
Under lång tid förlorade flercelliga eukaryoter konkurrenskamp prokaryota organismer, dock i slutet av Proterozoikum (800-600 miljoner år sedan) på grund av en kraftig förändring i förhållandena på jorden - en minskning av havsnivån, en ökning av syrekoncentrationen, en minskning av koncentrationen av karbonater i havsvatten , regelbundna kylningscykler - flercelliga eukaryoter fick fördelar jämfört med prokaryoter. Om det fram till den tiden bara fanns individuella flercelliga växter och, möjligen, svampar, så är djur också kända från det ögonblicket i jordens historia. Av de fauna som uppstod i slutet av Proterozoikum är Ediacaran och Vendian de bäst studerade. Det är vanligt att inkludera djur från den vendianska perioden i en speciell grupp av organismer eller att inkludera sådana typer som coelenterates, plattmaskar, leddjur etc. Ingen av dessa grupper har dock skelett, vilket kan tyda på frånvaron av rovdjur.
Livsutveckling under paleozoikum. Den paleozoiska eran, som varade i mer än 300 miljoner år, är indelad i sex perioder: kambrium, ordovicium, silur, devon, karbon (karbon) och perm.
V kambriska perioden land bestod av flera kontinenter, huvudsakligen belägna på södra halvklotet. De mest talrika fotosyntetiska organismerna under denna period var cyanobakterier och röda alger. Foraminifera och radiolarier levde i vattenpelaren. Ett stort antal skelettdjursorganismer förekommer i Kambrium, vilket framgår av många fossiler. Dessa organismer tillhörde cirka 100 typer av flercelliga djur, både moderna (svampar, coelenterater, maskar, leddjur, blötdjur) och utdöda, till exempel: det enorma rovdjuret anomalokaris och koloniala graptoliter som flöt i vattenpelaren eller var fästa på botten. Den torra marken förblev nästan obebodd i hela Kambrium, men markbildningsprocessen hade redan börjat av bakterier, svampar och möjligen lavar, och i slutet av perioden kom småborstar maskar och tusenfotingar ut på landet.
V Ordoviciumperiod havsnivån steg, vilket ledde till översvämningen av det kontinentala låglandet. De främsta producenterna under denna period var gröna, bruna och röda alger. Till skillnad från Kambrium, där reven byggde svampar, ersätts de i Ordovicium av korallpolyper. Snäckorna och bläckfiskar samt trilobiter (nu utdöda släktingar till spindeldjur). Under denna period spelades också in ackord för första gången, i synnerhet de käklösa. I slutet av Ordovicium inträffade en enorm utrotning, som förstörde cirka 35% av familjerna och mer än 50% av släktena av marina djur.
Silur kännetecknas av ökat bergsbyggande, vilket ledde till dränering av kontinentala plattformar. Den ledande rollen i den siluriska ryggradslösa faunan spelades av bläckfiskar, tagghudingar och gigantiska kräftdjur, medan det bland ryggradsdjur finns en mängd olika käklösa och fiskar. I slutet av perioden kom de första kärlväxterna, noshörningar och lycopoder, ut på land och började kolonisera de grunda vattnen och kusternas tidvattenzon. De första representanterna för arachnidklassen kom också ut på land.
V Devonperioden som ett resultat av landets uppgång bildades stora grunda vatten, som torkade upp och till och med frös igenom, eftersom klimatet blev ännu mer kontinentalt än i Silur. Haven domineras av koraller och tagghudingar, medan bläckfiskar representeras av spiralvridna ammoniter. Bland devonens ryggradsdjur nådde fiskarna sin storhetstid, och broskiga och beniga, såväl som lungor och korsfenade, kom att ersätta skalen. I slutet av perioden dyker de första groddjuren upp, som först levde i vatten.
I Mellandevonen dök de första skogarna av ormbunkar, lycopoder och åkerfräken upp på land, som var bebodda av maskar och många leddjur (tusenfotingar, spindlar, skorpioner, vinglösa insekter). I slutet av devon dök de första gymnospermerna upp. Assimileringen av mark av växter ledde till en minskning av vittringen och en ökning av jordbildningen. Konsolideringen av jordar ledde till uppkomsten av flodbäddar.
V Kolperiod land representerades av två kontinenter, åtskilda av ett hav, och klimatet blev märkbart varmare och fuktigare. I slutet av perioden skedde en liten landhöjning och klimatet ändrades till ett mer kontinentalt. Haven dominerades av foraminifer, koraller, tagghudingar, brosk och benig fisk, och sötvattenkroppar beboddes av musslor, kräftdjur och en mängd olika amfibier. I mitten av karbon uppstod små insektsätande reptiler och bevingade (kackerlackor, trollsländor) dök upp bland insekter.
Tropikerna kännetecknades av sumpiga skogar, dominerade av gigantiska åkerfräken, plonger och ormbunkar, vars döda rester sedan bildade avlagringar av kol. I mitten av perioden i den tempererade zonen, tack vare deras oberoende från vatten under befruktning och närvaron av sperma, började spridningen av gymnospermer.
Permperiod kännetecknas av sammanslagningen av alla kontinenter till en enda superkontinent Pangea, havens reträtt och klimatets kontinentitet stärkts i en sådan utsträckning att öknar bildades i Pangeas inre regioner. I slutet av perioden på land försvann trädormbunkar, åkerfränder och mossa nästan, och torkbeständiga gymnospermer tog den dominerande ställningen. Trots att stora groddjur fortfarande fortsatte att existera uppstod olika grupper av reptiler, inklusive stora växtätare och köttätande. I slutet av Perm inträffade den största utrotningen i livets historia, eftersom många grupper av koraller, trilobiter, de flesta bläckfiskar, fiskar (främst broskiga och korsfenade) och groddjur försvann. Den marina faunan förlorade 40–50 % av familjerna och cirka 70 % av släktena.
Livsutveckling i mesozoikum. Den mesozoiska eran varade i cirka 165 miljoner år och kännetecknades av landhöjning, intensivt bergsbyggande och en minskning av klimatets luftfuktighet. Den är indelad i tre perioder: trias, jura och krita.
I början Trias klimatet var torrt, men senare, på grund av höjningen av havsnivån, blev det fuktigare. Gymnospermer, ormbunkar och åkerfränder dominerade bland växterna, men de trädlevande formerna av sporbärande arter var nästan helt utrotade. Vissa koraller, ammoniter, nya grupper av foraminifer, musslor och tagghudingar har nått hög utveckling, samtidigt som mångfalden av broskfiskar har minskat, och grupper av benfiskar har också förändrats. Reptiler som dominerade land började bemästra vattenmiljön, som ichthyosaurs och plesiosaurs. Bland reptilerna i trias har krokodiler, tuataror och sköldpaddor överlevt till denna dag. I slutet av triasen dök dinosaurier, däggdjur och fåglar upp.
V Jurassic superkontinenten Pangea splittrades i flera mindre. Det mesta av jura var mycket fuktigt, och mot slutet blev klimatet torrare. Den dominerande gruppen av växter var gymnospermer, av vilka sequoia har överlevt från den tiden. Blötdjur (ammoniter och belemniter, musslor och snäckor), svampar, sjöborrar, brosk- och benfiskar blomstrade i haven. Stora groddjur dog nästan helt ut under juraperioden, men dök upp moderna grupper amfibier (svanslösa och svanslösa) och fjällande (ödlor och ormar), har mångfalden av däggdjur ökat. I slutet av perioden dök de möjliga förfäderna till de första fåglarna upp - Archaeopteryx. Alla ekosystem dominerades dock av reptiler - iktyosaurier och plesiosaurier, dinosaurier och flygande dinosaurier - pterosaurier.
Kritaperiod fick sitt namn i samband med att krita bildades i den tidens sedimentära bergarter. Över hela jorden, förutom polarområdena, rådde ett ihållande varmt och fuktigt klimat. Under denna period uppstod angiospermer och blev utbredda och ersatte gymnospermer, vilket ledde till en kraftig ökning av mångfalden av insekter. I haven dök, förutom blötdjur, teleostfiskar, plesiosaurier, ett stort antal foraminifer upp igen, vars skal bildade kalkavlagringar och dinosaurier dominerade på land. Fåglar bättre anpassade till luften började gradvis tränga undan de flygande dinosaurierna.
I slutet av perioden inträffade en global utrotning, som ett resultat av vilket ammoniter, belemniter, dinosaurier, pterosaurier och havsödlor, gamla grupper av fåglar och även några gymnospermer försvann. Cirka 16 % av familjerna och 50 % av djursläktena har försvunnit från jordens yta som helhet. Krisen i slutet av krita är förknippad med fallet av en stor meteorit i Mexikanska golfen, men det var troligen inte den enda orsaken till globala förändringar. Under det efterföljande köldknäppet överlevde bara små reptiler och varmblodiga däggdjur.
Utveckling av livet i kenozoikum. Den kenozoiska eran började för cirka 66 miljoner år sedan och fortsätter till denna dag. Det kännetecknas av dominansen av insekter, fåglar, däggdjur och angiospermer. Kenozoikum är indelat i tre perioder - paleogen, Neogene och Anthropogen - varav den sista är den kortaste i jordens historia.
Under den tidiga och mellersta paleogenen förblev klimatet varmt och fuktigt, i slutet av perioden blev det svalare och torrare. Angiospermer blev den dominerande gruppen av växter, men om det i början av perioden rådde vintergröna skogar, så uppträdde i slutet många lövskogar och stäpper bildades i torra zoner.
Bland fiskar intog benfiskar en dominerande ställning och antalet broskartade arter är obetydligt, trots deras betydande roll i saltvattenförekomster. På land, av reptilerna, överlevde bara fjäll, krokodiler och sköldpaddor, medan däggdjur ockuperade de flesta av sina ekologiska nischer. I mitten av perioden dök huvudorden av däggdjur upp, inklusive insektsätare, köttätare, pinnipeds, valar, klövdjur och primater. Isolering av kontinenter gjorde fauna och flora geografiskt mer mångsidig: Sydamerika och Australien blev centrum för utveckling av pungdjur och andra kontinenter - placenta däggdjur.
Neogen period. Jordytan i Neogen fick ett modernt utseende. Klimatet blev svalare och torrare. I Neogene var alla ordnar av moderna däggdjur redan bildade, och Hominid-familjen och Mankind-släktet uppstod i de afrikanska höljena. I slutet av perioden spred sig barrskogar i de cirkumpolära regionerna på kontinenterna, tundra dök upp och gräs ockuperade stäpparna i den tempererade zonen.
Kvartärperiod(antropogen) kännetecknas av periodiska förändringar av istider och uppvärmning. Under glaciärer täcktes höga breddgrader med glaciärer, havsnivån sjönk kraftigt, de tropiska och subtropiska bälten smalnade. I territorierna som gränsar till glaciärerna etablerades ett kallt och torrt klimat, vilket bidrog till bildandet av köldbeständiga grupper av djur - mammutar, jättehjortar, grottlejon etc. Den åtföljande glaciala minskningen av nivån på världshavet ledde till bildandet av landbroar mellan Asien och Nordamerika, Europa och de brittiska öarna etc. Djurvandringar ledde å ena sidan till ömsesidig berikning av flora och fauna, och å andra sidan till att utomjordingar förflyttade reliker t.ex. pungdjur och klövdjur i Sydamerika. Dessa processer påverkade dock inte Australien, som förblev isolerat.
I allmänhet har periodiska klimatförändringar lett till bildandet av en extremt rik artdiversitet som är karakteristisk för det nuvarande stadiet av biosfärens evolution, och även påverkat människans evolution. Under antropogenerna har flera arter av släktet Man bosatt sig från Afrika till Eurasien. För cirka 200 tusen år sedan dök arten Homo sapiens upp i Afrika, som efter en lång period av existens i Afrika för cirka 70 tusen år sedan gick till Eurasien och för cirka 35-40 tusen år sedan - till Amerika. Efter en period av samexistens med närbesläktade arter fördrev han dem och bosatte sig över hela världen. För cirka 10 tusen år sedan började mänsklig ekonomisk aktivitet i måttligt varma områden på jorden att påverka både planetens utseende (plöjning av mark, eldning av skogar, överbetning, ökenspridning etc.), och på fauna och flora p.g.a. minskningen av livsmiljöer deras livsmiljö och utrotning, och den antropogena faktorn kom in i bilden.
Mänskligt ursprung. Människan som art, hennes plats i den organiska världens system. Hypoteser om mänskligt ursprung. Drivkrafter och stadier av mänsklig evolution. Människoraser, deras genetiska förhållande. Människans biosociala natur. Social och naturlig miljö, mänsklig anpassning till den
Mänskligt ursprung
Inte ens för 100 år sedan trodde den överväldigande majoriteten av människor på planeten att människan kunde härstamma från så "små respektabla" djur som apor. I en diskussion med en av försvararna av den darwinistiska evolutionsteorin frågade professor Thomas Huxley, hans ivrige motståndare, biskopen av Oxford, Samuel Wilberforce, som förlitade sig på religiösa dogmer, honom till och med om han ansåg sig vara kopplad till apans förfäder genom sin farfar. eller mormor.
Ändå uttrycktes tankar om det evolutionära ursprunget av antika filosofer, och den store svenske taxonomen K. Linné gav på 1700-talet, utifrån en uppsättning tecken, ett artnamn till människan Homo sapiens L.(Homo sapiens) och tog honom tillsammans med aporna till en ordning - primaterna. JB Lamarck stöttade C. Linnaeus och trodde att människan till och med hade gemensamma förfäder med moderna apor, men någon gång i sin historia härstammade hon från ett träd, vilket var en av anledningarna till att människan bildades som art.
Charles Darwin ignorerade inte heller denna fråga och publicerade på 70-talet av XIX-talet verken "The Origin of Man and Sexual Selection" och "On the Expression of Emotions in Animals and Man", där han gav inte mindre övertygande bevis för människans och apornas gemensamma ursprung än den tyske forskaren E. Haeckel ("Skapelsens naturhistoria", 1868; "Anthropogenesis, or the History of the Origin of Man", 1874), som till och med sammanställde djurets genealogi. rike. Dessa studier rörde dock bara den biologiska sidan av bildningen av människan som art, medan de sociala aspekterna avslöjades av den klassiska historiska materialismen - den tyske filosofen F. Engels.
För närvarande, ursprunget och utvecklingen av människan som en biologisk art, såväl som mångfalden av populationer modern man och lagarna för deras interaktion studeras av vetenskapen antropologi.
Människan som art, hennes plats i den organiska världens system
Homo sapiens ( Homo sapiens) som en biologisk art tillhör djurriket, underriket flercelliga. Närvaron i processen av embryonal utveckling av notokordet, gälslitsar i svalget, neuralröret och bilateral symmetri gör det möjligt att klassificera det som kordat, medan utvecklingen av ryggraden, närvaron av två par lemmar och placeringen av hjärtat på den ventrala sidan av kroppen indikerar dess förhållande till andra representanter för ryggradsdjurssubtypen.
Matning av spädbarn med mjölk som utsöndras av bröstkörtlarna, varmblodighet, fyrkammarhjärta, förekomst av hår på kroppens yta, sju kotor i halsryggraden ryggraden, munnens vestibul, alveolära tänder och bytet av mjölktänder till permanenta är tecken på klassen däggdjur, och embryots intrauterina utveckling och dess förbindelse med moderns kropp genom moderkakan är en underklass av placenta. .
Mer frekventa tecken, såsom att greppa lemmar med motsatta tumme och naglar, utveckling av nyckelbenen, framåtblickande ögon, en ökning av storleken på skallen och hjärnan, och närvaron av alla grupper av tänder (framtänder, hörntänder och molarer) lämna inget tvivel om att hans plats är i ordningen primater.
Den betydande utvecklingen av hjärnan och härmarmusklerna, såväl som tändernas strukturella egenskaper, gör det möjligt att klassificera en person som en underordning av högre primater, eller apor.
Frånvaron av en svans, närvaron av böjningar av ryggraden, utvecklingen av hjärnhalvorna i framhjärnan, täckt med en bark med många spår och veck, närvaron av överläppen och hårfästets uttunning ger anledning att placera den bland representanterna för familjen av högre smalnäsa eller människoapor.
Men även de mest välorganiserade mänskliga aporna kännetecknas av en kraftig ökning av hjärnans volym, upprätt hållning, ett brett bäcken, en utskjutande haka, artikulerat tal och närvaron av 46 kromosomer i karyotypen och bestämmer dess tillhörighet till karyotypen. släktet Man.
Användningen av de övre extremiteterna för arbete, tillverkning av verktyg, abstrakt tänkande, kollektiv aktivitet och utveckling baserad på mer sociala än biologiska lagar, är de specifika egenskaperna hos Homo sapiens.
Alla moderna människor tillhör en art - Homo sapiens ( Homo sapiens), och underarter H. sapiens sapiens... Denna art är en uppsättning populationer som ger fertil avkomma när de korsas. Trots en ganska betydande variation av morfofysiologiska karaktärer är de inte bevis på en högre eller lägre grad av organisation av vissa grupper av människor - de är alla på samma utvecklingsnivå.
I vår tid har ett tillräckligt antal vetenskapliga fakta redan samlats in för att skapa människan som art i evolutionsprocessen - antropogenes... Det specifika förloppet av antropogenesen är ännu inte helt klarlagt, men tack vare nya paleontologiska rön och moderna forskningsmetoder kan man hoppas att en tydlig bild snart nog kommer fram.
Hypoteser om mänskligt ursprung
Om vi inte tar hänsyn till hypoteserna om människans gudomliga skapelse och hennes penetration från andra planeter, som inte är relaterade till biologins område, så leder alla mer eller mindre konsekventa hypoteser om människans ursprung honom till gemensamma förfäder med moderna primater.
Så, hypotes om ursprunget till människan från den gamla tropiska primaten tarsier, eller Tarzials hypotes, formulerad av den engelske biologen F. Wood Jones 1929, förlitar sig på likheten mellan proportionerna av de mänskliga och tarsier kropparna, egenskaperna hos hårfästet, förkortningen av ansiktsdelen av skallen hos den senare, etc. Emellertid, skillnaderna i dessa organismers struktur och vitala aktivitet är så stora att de inte har vunnit allmän acceptans.
Med antropoida apor har människor till och med för många likheter. Så, förutom de anatomiska och morfologiska egenskaperna som redan nämnts ovan, bör uppmärksamhet ägnas åt deras postembryonala utveckling. Till exempel har små schimpanser mycket glesare hår, ett mycket större förhållande mellan hjärna och kropp och ett något bredare utbud av bakben än vuxna. Även sexuell mognad hos människoapor sker mycket senare än hos representanter för andra däggdjursordningar med liknande kroppsstorlekar.
Under loppet av cytogenetiska studier avslöjades det att en av de mänskliga kromosomerna bildades som ett resultat av sammansmältningen av kromosomer av två olika par som finns i karyotypen av apor, och detta förklarar skillnaden i antalet kromosomer (i människor, 2n = 46, och hos stora apor, 2n = 48 ), och är också ett annat bevis på förhållandet mellan dessa organismer.
Likheten mellan människor och människoapor är också mycket stor i molekylär-biokemiska data, eftersom människor och schimpanser har samma AB0- och rhesus-blodgruppsproteiner, många enzymer och aminosyrasekvenserna i hemoglobinkedjorna har endast 1,6 % skillnader, medan med andra apor denna diskrepans något mer. Och på genetisk nivå är skillnaderna i nukleotidsekvenserna i DNA mellan dessa två organismer mindre än 1%. Om vi tar hänsyn till den genomsnittliga utvecklingshastigheten för sådana proteiner i besläktade grupper av organismer, kan det fastställas att mänskliga förfäder separerade från andra grupper av primater för cirka 6-8 miljoner år sedan.
Apornas beteende liknar på många sätt människors, eftersom de lever i grupper där sociala roller är tydligt fördelade. Gemensamt skydd, ömsesidig hjälp och jakt är inte de enda målen för att skapa en grupp, eftersom aporna inom den känner tillgivenhet för varandra, uttrycker det på alla möjliga sätt och reagerar känslomässigt på olika stimuli. Dessutom sker i grupp ett erfarenhetsutbyte mellan individer.
Människans likhet med andra primater, särskilt de högre smalnosade aporna, finns alltså på olika nivåer av biologisk organisation, och skillnaderna mellan människan som art bestäms till stor del av egenskaperna hos denna grupp av däggdjur.
Hypoteserna om polycentrism och monocentrism tillhör den grupp av hypoteser som inte ifrågasätter människans ursprung från gemensamma förfäder med moderna apor.
Första position polycentrisk hypotesär uppkomsten och parallella evolutionen av den moderna arten av människan i flera regioner av världen samtidigt från olika former av forntida eller till och med den äldsta människan, men detta motsäger huvudbestämmelserna i den syntetiska evolutionsteorin.
Hypoteser om den moderna människans gemensamma ursprung, tvärtom, postulerar människans uppkomst på ett ställe, men skiljer sig åt var det hände. Så, hypotes om extratropiskt mänskligt ursprung bygger på det faktum att endast de hårda klimatförhållandena på de höga breddgraderna i Eurasien skulle kunna bidra till "humanisering" av apor. Upptäckten på Yakutias territorium av platser från tiden för den äldsta paleolitiska tiden - Deering-kulturen, vittnade till dess fördel, men senare fastställdes det att åldern för dessa fynd inte är 1,8–3,2 miljoner år, utan 260–370 år. tusen år. Denna hypotes är alltså inte heller tillräckligt bekräftad.
Den största mängden bevis samlas för närvarande in specifikt till förmån för hypotes om mänsklig afrikansk härkomst, men det saknar inte brister, som är utformade för att ta hänsyn till ett komplex bred monocentrismhypotes, som kombinerar argumenten för hypoteserna om polycentrism och monocentrism.
Drivkrafter och stadier i mänsklig evolution
Till skillnad från andra representanter för djurvärlden exponerades människan under sin evolution inte bara för biologiska evolutionsfaktorer utan också sociala, vilket bidrog till uppkomsten av en art av kvalitativt nya varelser med biosociala egenskaper. Sociala faktorer ledde till ett genombrott i en fundamentalt ny adaptiv miljö, vilket gav enorma fördelar för mänskliga befolkningars överlevnad och kraftigt accelererade dess utvecklingshastighet.
Biologiska faktorer av evolution, som spelar en viss roll i antropogenesen till denna dag, är ärftlig variation, såväl som flödet av gener som tillhandahåller det primära materialet för naturligt urval. Samtidigt har isolering, befolkningsvågor och gendrift nästan helt förlorat sin betydelse till följd av vetenskapliga och tekniska framsteg. Detta ger anledning för vissa forskare att tro att i framtiden kommer även minimala skillnader mellan representanter för olika raser att försvinna på grund av deras blandning.
Eftersom förändringen i miljöförhållandena tvingade människans förfäder att gå ner från träd till öppna utrymmen och gå över till att gå på två lemmar, användes de släppta övre extremiteterna av dem för att bära mat och barn, samt för att tillverka och använda verktyg. Ett sådant verktyg kan dock endast göras om det finns en tydlig uppfattning om det slutliga resultatet - bilden av objektet, därför utvecklades också abstrakt tänkande. Det är välkänt att komplexa rörelser och tankeprocessen är nödvändiga för utvecklingen av vissa områden i hjärnbarken, vilket har hänt under evolutionsprocessen. Det är dock omöjligt att ärva sådana kunskaper och färdigheter, de kan bara överföras från en individ till en annan under den senares liv, vilket resulterade i skapandet av en speciell form av kommunikation - artikulerat tal.
Alltså till sociala faktorer evolution bör tillskrivas arbetsaktivitet mänskligt, abstrakt tänkande och artikulerat tal. Uttrycken av den primitiva mannens altruism, som tog hand om barn, kvinnor och äldre, bör inte heller förkastas.
En persons arbetsaktivitet påverkade inte bara utseendet på sig själv utan tillät till en början att delvis lindra existensvillkoren genom att använda eld, tillverka kläder, bygga bostäder och i framtiden aktivt förändra dem genom avskogning, plöjning land, etc. har okontrollerad ekonomisk aktivitet ställt mänskligheten inför hotet om en global katastrof till följd av jorderosion, uttorkning av sötvattenreservoarer, förstörelse av ozonskärmen, vilket i sin tur kan öka trycket på biologiska faktorer av evolution.
Driopithecus, som levde för cirka 24 miljoner år sedan, var troligen den gemensamma förfadern till människor och människoapor. Trots att han klättrade i träd och sprang på alla fyra lemmar kunde han gå på två ben och bära mat i händerna. Den fullständiga separationen av de högre aporna och linjen som leder till människor ägde rum för cirka 5-8 miljoner år sedan.

Australopithecus... Från Dryopithecus fanns det tydligen ett släkte ardipithecus, som bildades för över 4 miljoner år sedan på Afrikas savanner som ett resultat av en köldknäpp och skogarnas reträtt, vilket tvingade dessa apor att röra sig på bakbenen. Detta lilla djur gav uppenbarligen upphov till ett ganska talrikt släkte. Australopithecus("Södra apa").
Australopithecus dök upp för cirka 4 miljoner år sedan och levde i de afrikanska savannerna och torra skogarna, där fördelarna med tvåfota rörelser manifesterades fullt ut. Från Australopithecus gick två grenar - stora växtätare med kraftiga käkar parantroper och mindre och mindre specialiserade människor... Under loppet av en viss tid utvecklades dessa två släkten parallellt, vilket i synnerhet manifesterade sig i en ökning av hjärnans volym och komplikationen av de använda verktygen. Egenskaperna i vår familj är tillverkningen av stenverktyg (parantroper som endast används ben) och en relativt stor hjärna.
De första representanterna för släktet Man dök upp för cirka 2,4 miljoner år sedan. De tillhörde typen av en skicklig man (Homo habilis) och var korta varelser (cirka 1,5 m) med en hjärnvolym på cirka 670 cm 3. De använde grova stenverktyg. Tydligen hade representanterna för denna art välutvecklade ansiktsuttryck och rudimentärt tal. En skicklig man lämnade den historiska scenen för cirka 1,5 miljoner år sedan, vilket gav upphov till följande arter - en uträtad person.
Man rätade ut sig (H. erectus) som en biologisk art som bildades i Afrika för cirka 1,6 miljoner år sedan och existerade i 1,5 miljoner år och spred sig snabbt över stora territorier i Asien och Europa. En representant för denna art från ön Java beskrevs vid en tidpunkt som pithecanthropus("Monkey man") som upptäcktes i Kina fick sitt namn sinanthropus, medan deras europeiska "motsvarighet" är Heidelberg man.
Alla dessa former kallas också arkantroper(av de äldsta människorna). En uträtad man kännetecknades av en låg panna, stora ögonbrynskanter och en lutande bakre haka, hjärnvolymen var 900–1200 cm 3. En uträtad persons kropp och lemmar liknade en modern persons. Utan tvekan använde representanter för denna familj eld och gjorde tvåeggade kotletter. Som de senaste fynden har visat, behärskade denna art till och med navigering, eftersom dess ättlingar hittades på avlägsna öar.
Paleoanthropus. Cirka 200 tusen år sedan från Heidelberg kom mannen neandertalman (H. neandertalensis) som tillhör paleoanthropus(till forntida människor), som levde i Europa och västra Asien för 200-28 tusen år sedan, inklusive under glaciärernas epok. De var starka, tillräckligt fysiskt starka och tåliga människor med en stor hjärnvolym (till och med större än en modern persons). De hade artikulerat tal, tillverkade sofistikerade verktyg och kläder, begravde sina döda och kanske till och med hade några konstverk. Neandertalarna var inte förfäder till Homo sapiens, denna grupp utvecklades parallellt. Deras utrotning är förknippat med försvinnandet av mammutfaunan efter den senaste nedisningen, och är möjligen också resultatet av konkurrerande förskjutning av vår art.
Det äldsta fyndet av en representant Homo sapiensär 195 tusen år gammal och kommer från Afrika. Troligtvis är den moderna människans förfäder inte neandertalare, utan någon form av archantropus, till exempel Heidelberg-människan.
Neoantropus. För cirka 60 tusen år sedan, som ett resultat av okända händelser, dog vår art nästan ut, så alla följa människorär ättlingar till en liten grupp som endast bestod av några dussin individer. Genom att övervinna denna kris började vår art bosätta sig i Afrika och Eurasien. Den skiljer sig från andra arter i en slankare kroppsbyggnad, en högre reproduktionshastighet, aggressivitet och, naturligtvis, det mest komplexa och mest flexibla beteendet. Människor av den moderna typen som bebodde Europa för 40 tusen år sedan kallas Cro-Magnons och hänvisa till neoantroper(till moderna människor). De skilde sig inte biologiskt från moderna människor: höjd 170–180 cm, hjärnvolym cirka 1600 cm 3. Cro-Magnons utvecklade konst och religion, de domesticerade många arter av vilda djur och domesticerade många arter av växter. Moderna människor härstammar från Cro-Magnon.
Människoraser, deras genetiska förhållande
Med vidarebosättning av mänskligheten på planeten mellan olika grupper människor, det fanns vissa avvikelser när det gäller hudfärg, ansiktsdrag, hårets karaktär samt frekvensen av förekomsten av vissa biokemiska egenskaper. Helheten av sådana ärftliga egenskaper kännetecknar en grupp individer av samma art, vars skillnader är mindre betydande än underarten - lopp.
Studiet och klassificeringen av raser kompliceras av bristen på tydliga gränser mellan dem. Hela den moderna mänskligheten tillhör en art, inom vilken tre stora raser urskiljs: Australo-Negroid (svart), Caucasoid (vit) och Mongoloid (gul). Var och en av dem är uppdelad i små raser. Skillnader mellan raser beror på egenskaper som hudfärg, hår, näsform, läppar, etc.
australisk negroid, eller ekvatoriallopp kännetecknas av mörk hudfärg, vågigt eller lockigt hår, bred och något utstående näsa, tvärgående näsborrar, tjocka läppar och ett antal kraniala tecken. Kaukasoid, eller eurasisk ras kännetecknas av ljus eller mörk hud, rakt eller vågigt mjukt hår, bra utveckling hår i ansiktet på män (skägg och mustasch), en smal utstående näsa, tunna läppar och ett antal kraniala tecken. Mongoloid(asiatisk amerikan) lopp kännetecknas av mörk eller ljus hud, ofta grovt hår, medelbredd på näsan och läpparna, tillplattad ansikte, starkt utskjutande kindben, relativt stor ansiktsstorlek, märkbar utveckling av "det tredje århundradet".
Dessa tre raser skiljer sig också åt i avveckling. Före den europeiska koloniseringens era var den australisk-negroida rasen utbredd i den gamla världen söder om Kräftans vändkrets; Kaukasoid ras - i Europa, Nordafrika, Västasien och norra Indien; Mongoloid ras - i sydöstra, norra, centrala och östra Asien, Indonesien, Nord- och Sydamerika.
Skillnaderna mellan raser gäller dock endast mindre egenskaper som har en adaptiv betydelse. Så huden på negroider bränns vid en tio gånger högre dos av ultraviolett strålning än huden på kaukasier, men kaukasier lider mindre av rakitis på höga breddgrader, där det kan finnas en brist på ultraviolett strålning som är nödvändig för bildandet av vitamin D.
Tidigare försökte vissa människor bevisa perfektionen hos en av raserna för att få en moralisk fördel gentemot andra. För närvarande är det tydligt att rasegenskaper endast återspeglar de olika historiska vägarna för grupper av människor, men är inte på något sätt kopplade till fördelen eller den biologiska efterblivenheten hos en eller annan grupp. Människoraser är mindre väldefinierade än andra djurs underarter och raser, och kan inte på något sätt jämföras till exempel med raser av husdjur (som är resultatet av målmedvetet urval). Biomedicinsk forskning visar att effekterna av äktenskap mellan olika raser beror på mannens och kvinnans individuella egenskaper och inte på deras ras. Därför är alla förbud mot rasäktenskap eller viss vidskepelse ovetenskapliga och omänskliga.
Mer specifika än raser är grupper av människor nationaliteter- historiskt bildade språkliga, territoriella, ekonomiska och kulturella gemenskaper av människor. Befolkningen i ett visst land bildar dess folk. Med samverkan mellan många nationaliteter kan en nation uppstå som en del av ett folk. Nu på jorden finns det inga "rena" raser, och varje tillräckligt stort folk representeras av människor som tillhör olika raser.
Människans biosociala natur
Utan tvekan måste människor som art vara under trycket av evolutionära faktorer som mutagenes, befolkningsvågor och isolering. Men när det mänskliga samhället mognar försvagas vissa av dem, medan andra tvärtom intensifieras, eftersom det nästan inte finns några isolerade mänskliga populationer kvar på planeten, fångad av globaliseringsprocesserna, där närbesläktade korsningar utförs, och antalet populationer i sig är inte föremål för kraftiga fluktuationer. Följaktligen spelar den drivande faktorn för evolutionen - naturligt urval - tack vare framgången för medicin, inte längre den roll i mänskliga populationer som är karakteristisk för den i populationer av andra organismer.
Tyvärr leder försvagningen av selektionstrycket till en ökning av frekvensen av ärftliga sjukdomar i populationer. Till exempel i industrialiserade länder lider upp till 5 % av befolkningen av färgblindhet (färgblindhet), medan denna siffra i mindre utvecklade länder är upp till 2 %. Negativa konsekvenser detta fenomen kan övervinnas genom förebyggande åtgärder och framsteg inom vetenskapsområden som genterapi.
Detta betyder dock inte att mänsklig evolution är över, eftersom det naturliga urvalet fortsätter att fungera och eliminerar t.ex. könsceller och individer med ogynnsamma genkombinationer även vid de proembryonala och embryonala perioder ontogenes, såväl som resistens mot patogener av olika sjukdomar. Dessutom tillhandahåller materialet för naturligt urval inte bara mutationsprocessen, utan också ackumuleringen av kunskap, förmågan att lära, uppfattningen av kultur och andra egenskaper som kan överföras från person till person. Till skillnad från genetisk information överförs den erfarenhet som ackumuleras i processen för individuell utveckling både från föräldrar till avkomma och i motsatt riktning. Och konkurrens uppstår redan mellan samhällen som är kulturellt olika. Denna form av evolution, som uteslutande är speciell för människor, kallas kulturell, eller social evolution.
Ändå utesluter inte kulturell evolution biologisk evolution, eftersom den blev möjlig endast som ett resultat av bildningen mänsklig hjärna, och den mänskliga biologin i sig bestäms för närvarande av kulturell evolution, eftersom i frånvaro av samhället och en mängd olika rörelser i hjärnan bildas inte vissa zoner.
Således har en person en biosocial natur, som lämnar ett avtryck på manifestationen av biologiska, inklusive genetiska, lagar som styr hans individuella och evolutionära utveckling.
Social och naturlig miljö, mänsklig anpassning till den
Under social miljö först och främst förstå de sociala materiella och andliga förutsättningarna för hans existens och verksamhet kring en person. Förutom det ekonomiska systemet, sociala relationer, socialt medvetande och kultur, inkluderar det också den närmaste omgivningen för en person - familjen, arbets- och studentkollektiven, såväl som andra grupper. Omgivningen har å ena sidan ett avgörande inflytande på personlighetens bildning och utveckling, och å andra sidan förändras den själv under människans inflytande, vilket medför nya förändringar hos människor osv.
Anpassning av individer eller deras grupper till den sociala miljön för att förverkliga deras egna behov, intressen, livsmål och inkluderar anpassning till förutsättningarna och karaktären av studier, arbete, mellanmänskliga relationer, ekologisk och kulturell miljö, villkor för fritid och vardagsliv , såväl som deras aktiva förändring för att möta deras behov. En stor roll i detta spelas genom att förändra sig själv, sina motiv, värderingar, behov, beteende osv.
Informationsmängder och känslomässiga upplevelser i moderna samhälletär ofta den främsta orsaken till stress, som kan övervinnas med hjälp av tydlig självorganisering, fysisk träning och autoträning. I vissa, särskilt svåra fall, krävs att man konsulterar en psykoterapeut. Ett försök att hitta glömska av dessa problem i att äta för mycket, röka, dricka alkohol och andra dåliga vanor leder inte till det önskade resultatet, utan förvärrar bara kroppens tillstånd.
Den naturliga miljön har inte mindre inflytande på människor, trots att en person i cirka 10 tusen år har försökt att skapa en bekväm konstgjord miljö för sig själv. Så att klättra till en avsevärd höjd på grund av en minskning av koncentrationen av syre i luften leder till en ökning av antalet röda blodkroppar i blodet, ökad andning och hjärtslag, och långvarig exponering för den öppna solen bidrar till ökad hud pigmentering - garvning. De angivna förändringarna passar dock in i reaktionshastigheten och ärvs inte. Ändå kan folk som har levt under sådana förhållanden under lång tid ha vissa anpassningar. Så bland nordliga folk har näsbihålorna en mycket större volym för att värma luften, och storleken på de utskjutande delarna av kroppen minskar för att minska värmeförlusten. Afrikaner kännetecknas av mörkare hudfärg och lockigt hår, eftersom pigmentet melanin skyddar kroppens organ från penetration av skadliga ultravioletta strålar, och hårlocket har värmeisolerande egenskaper. Européernas ljusa ögon är en anpassning för en skarpare uppfattning av visuell information i skymning och dimma, och den mongoloida ögonformen är resultatet av naturligt urval för inverkan av vindar och dammstormar.
Dessa förändringar tar århundraden och årtusenden, men livet i ett civiliserat samhälle innebär vissa förändringar. Så, en minskning av fysisk aktivitet leder till en ljusning av skelettet och en minskning av dess styrka, en minskning muskelmassa... Låg rörlighet, ett överskott av kaloririk mat, stress medför en ökning av antalet överviktiga personer, och högvärdig proteinnäring och fortsättningen av dagsljus med hjälp av artificiell belysning bidrar till acceleration - acceleration av tillväxt och pubertet, en ökning av kroppsstorlek.
Alternativ 1.
Charles Darwins främsta förtjänst i utvecklingen av biologi är:
1) utveckla urvalsmetoder 2) identifiera evolutionens drivkrafter
3) skapandet av de vetenskapliga grunderna för taxonomi, 4) studiet av paleontologiska fynd.
Som ett resultat av samspelet mellan evolutionens drivkrafter inträffar följande:
1) fluktuation i antalet populationer 2) bildandet av nya arter i naturen
3) mutationsprocess 4) isolering av populationer.
Vilka är orsakerna till kampen för tillvaron?
1) individernas variation i befolkningen
2) begränsade resurser i miljön och intensiv reproduktion av individer
3) naturkatastrofer
4) bristen på anpassning av individer till miljön.
Vilken variation är materialet för naturligt urval?
1) säsongsbetonad 2) mutationell 3) bestämd 4) fenotypisk.
Nyttiga mutationer sprids genom befolkningen på grund av:
1) förflyttning av individer 2) fri korsning
3) fysiologisk isolering 4) miljöisolering.
Naturligt urval är:
1) komplexa relationer mellan organismer och den livlösa naturen
2) processen att bevara individer med fördelaktiga ärftliga förändringar
3) processen för bildandet av nya arter i naturen
4) processen för befolkningstillväxt.
I riktning mot anpassning av organismer till miljön verkar följande:
1) metodiskt urval 2) naturligt urval
3) ärftlig föränderlighet 4) kampen för tillvaron.
Urvalet av individer med egenskaper som avviker från medelvärdet kallas:
1) körning 2) metodisk 3) stabiliserande 4) massiv.
Vad är gendrift?
1) slumpmässig förändring i frekvensen av förekomsten av deras alleler i populationen
2) förflyttning av individer från en befolkning till en annan
Växter och djurs anpassningsförmåga till deras livsmiljö är resultatet:
1) individers önskan om självförbättring 2) mänskliga aktiviteter
3) modifieringsvariabilitet 4) samverkan mellan evolutionens drivkrafter.
Växters anpassningsförmåga till insektspollinering manifesteras i:
1) bildandet av en stor mängd pollen 2) förlängningen av filament
3) tidig vårblomning 4) närvaron av nektar i blommorna, en ljus krans.
Hur är den vita harens anpassningsförmåga att skydda sig mot rovdjur på vintern?
1) närvaron av en konstant kroppstemperatur 2) förändring av hårfästet
3) förekomsten av viloläge 4) förmågan att snabbt röra sig i snön.
Embryologiska bevis för evolution inkluderar:
1) cellstrukturen hos organismer
2) förekomsten av liknande organsystem hos ryggradsdjur
3) likheten mellan embryon från ryggradsdjur
4) likheten mellan livsprocesser hos djur.
Förhållandet mellan ryggradslösa djur och ryggradsdjur kan bevisas genom att undersöka strukturen:
1) broskfisk 2) annelider 3) benfisk 4) lansett.
jag ... Urvalskriterium Urvalstyp
A) bevarar individer med användbara under dessa förhållanden 1) naturliga
miljöförändringar.
B) leder till skapandet av nya djurraser 2) konstgjorda
och växtsorter.
C) främjar skapandet av organismer med nödvändiga
en person med ärftliga förändringar.
D) manifesterar sig inom befolkningen och mellan populationer
av samma art i naturen.
E) verkar i naturen i miljontals år.
E) leder till bildandet av nya arter och bildandet
anpassningsförmåga till miljön.
G) utförs av en person.
II ... Kriterium för funktionstyp
A) kroppen är täckt med en mantel. 1) morfologiska
B) skalet har två ventiler. 2) ekologiskt
C) lever i sötvatten.
D) cirkulationssystemet är inte stängt.
E) livnär sig på vattenlevande mikroorganismer.
E) larven utvecklas i vatten.
III. Organism Evolutionens riktning
A) strutsemu 1) biologiska framsteg
B) gråråtta 2) biologisk regression
C) husmus
Blågrön (cyanobakterier)
E) örn kungsörn
E) Ussuri tiger
Uppgift 3. Fastställ sekvensen för utseendet av aromorfoser i utvecklingen av ryggradsdjur:
A) tvåkammar fiskhjärta
B) utveckling av barnet i däggdjurets livmoder
C) inre befruktning hos reptiler
D) lungandning hos amfibier
Testa i biologi. Årskurs 11.
"Charles Darwins doktrin om evolutionens drivkrafter och resultat.
Syntetisk evolutionsteori".
Alternativ 2.
Uppgift 1. Välj rätt svar:
Vad är ärftlighet, föränderlighet, kamp för tillvaron och naturligt urval?
1) den levande naturens egenskaper 2) evolutionens resultat
3) evolutionens drivkrafter 4) evolutionens huvudriktningar.
Evolutionens drivkrafter inkluderar:
1) mångfald av arter 2) kamp för tillvaron
3) artbildning 4) kondition.
Vilken betydelse har kampen för existens i evolutionen?
1) bevarande av individer främst med användbara förändringar
2) bevarande av individer med eventuella ärftliga förändringar
3) skapa material för urval
4) förvärring av relationer mellan individer.
Vad ligger vinsten i naturlig befolkning mutationsprocess?
1) öka effektiviteten av naturligt urval
2) öka intensiteten i cirkulationen av ämnen
3) öka antalet individer, 4) förbättra självregleringen.
Vilken roll spelar recessiva mutationer i evolutionen?
1) visas i den första generationen
2) är en dold reserv av ärftlig variation
3) vanligtvis skadligt för kroppen 4) påverka generna i kroppsceller, inte könsceller.
Bildandet av nya arter i naturen sker som ett resultat av:
1) individers önskan om självförbättring
2) mänskligt bevarande av individer med fördelaktiga ärftliga förändringar
3) 0 bevarande av individer med fördelaktiga ärftliga förändringar av människor
4) fenotypisk.
bevarande genom naturligt urval av individer med fördelaktiga ärftliga förändringar.
4) bevarande genom naturligt urval av individer med en mängd icke-ärftliga förändringar.
Vad är namnet på urval, där individer med en genomsnittlig norm för en indikator på en egenskap bevaras i en population?
1) stabiliserande 2) körning 3) konstgjord 4) metodisk.
Vilka är konsekvenserna av handlingen att stabilisera urvalet?
1) bevarande av gamla arter 2) förändring i reaktionshastigheten
3) uppkomsten av nya arter, 4) bevarandet av individer med förändrade egenskaper.
Evolutionsfaktorn, vars grund är uppkomsten av hinder för individers fria korsning, kallas:
1) modifiering 2) naturligt urval
3) isolering 4) befolkningsvågor.
Vilka är orsakerna till mångfalden av arter i naturen?
1) säsongsmässiga förändringar i ekosystemet 2) organismernas anpassningsförmåga till miljön
3) ärftlighetsvariabilitet och naturligt urval
4) modifieringsvariabilitet och artificiellt urval.
Djurens anpassningsförmåga till säsongsmässiga förändringar i naturen är:
1) jaga hajar, rockor 2) flyttfåglar
3) nattlig aktivitet hos fladdermöss, 4) rörelse av blötdjursskalventilerna.
Vilken typ av enheter tillskrivs nyckelpigornas ljusa färger?
1) mimik 2) förklädnad
3) skyddsfärgning 4) varningsfärgning.
Ge ett exempel på paleontologiska bevis för evolution:
1) avtryck av Archaeopteryx 2) arternas mångfald av organismer
3) fiskens anpassningsförmåga till liv på olika djup, 4) förekomsten av skal i blötdjur.
Organ som har förlorat sin ursprungliga funktion under evolutionen kallas:
1) atavismer 2) rudiment 3) homologa 4) liknande.
Uppgift 2. Ställ in korrespondensen:
jag. Bevis för Evolution Statement
A) mänsklig ontogenes, som schimpanser, 1) embryologisk
börjar med en zygot.
B) en fågels vinge och en mullvads tass är homologa 2) jämförande anatomiska
C) i en flock av hästar, är utseendet på tretåiga möjliga
D) förekomsten av gälskåror i embryot hos ett däggdjur.
D) alla ryggradsdjur i individuell utveckling passera
stadier av blastula, gastrula, neurula.
II. Kriterium för funktionstyp
A) larven lever i vatten 1) morfologisk
B) kroppen är tillplattad. 2) ekologiskt
D) livnär sig på värdens vävnader.
D) har sugkoppar.
E) matsmältningssystemet har en munöppning.
III. Karakteristisk Miljöfaktor
A) konstantheten hos atmosfärens gassammansättning. 1) biotisk
B) ändra tjockleken på ozonskärmen. 2) abiotisk
C) förändring av luftfuktigheten.
D) förändring av antalet konsumenter.
E) förändring av antalet producenter.
Uppgift 3. Fastställ sekvensen av processerna för embryonal utveckling av ryggradsdjur:
A) bildandet av blastomerer under klyvningen av zygoten
B) läggning av embryots embryonala organ
C) sammansmältning av ägg och spermier och bildandet av en zygot
D) utveckling av neurala plattan
E) bildandet av groddlager
Provarbete i biologi. Årskurs 11.
"Charles Darwins doktrin om evolutionens drivkrafter och resultat.
Syntetisk evolutionsteori".
Svar. Alternativ 1.
Övning 1.
Uppgift 2.
III.
Uppgift 3.
Alternativ 2.
Övning 1.
III.







